First Person: Neuro-Cognitive Notes on the Self in Life and in Fiction
by
William L. Benzon
August 21, 2000
We can think of the self as the result of interaction between subcortical systems for regulating the global brain state and largely cortical systems for representing the current body state and autobiography. The personal pronoun system is at the interface between the cortical and subcortical systems. By constructing a network model for the pronoun system that is grounded in basic machinery for social interaction we show how the pronoun system allows speakers to achieve self-reference and how this capacity engenders the illusion of a unified self. The same model allows us to see that there is no essential difference between reliving incidents from one's own past and giving life to imaginary characters in ritual and in literary works. Such imaginative experience may play a role in maintaining the coherence of the self through different emotions.
|
Cogito ergo sum. --Rene Descartes It is in and through language that man constitutes himself as a subject, because language alone establishes the concept of "ego" in reality, in its reality which is that of the being. --Emile Benveniste |
At the beginning of the chapter on "The Consciousness of Self" in his magisterial The Principles of Psychology William James asserts that:
In its widest possible sense, however, a man's Self is the sum total of all that he CAN call his, not only his body and his psychic powers, but his clothes and his house, his wife and children, his ancestors and friends, his reputation and works, his lands and horses, and yacht and bank-account. All these things give him the same emotions. If they wax and prosper, he feels triumphant; if they dwindle and die away, he feels cast down . . .
Accepting the truth of these statements, one can easily conjure up a host of mysteries by asking just where in the brain one is to locate the Self that has this heterogeneous collection of appendages. One of the deepest mysteries, of course, stems from the fact that that very brain, in all its intricate detail of structure and dance of process, is itself but one of those appendages. It is but "my brain" and it is but a part of "my body" on behalf of which it enacts "my mind." What is this Self that can have a brain, a body, and a mind?
A common answer is that that Self is but an illusion, a ghost in the machine. Modern neuroscience presents us with a brain that has many parts intricately organized in many circuits on many levels. But no one component is supposed to be in charge of the whole operation (see e.g. Bownds 1999, 291ff.). If neuroscience thus relieves us of the burden of locating this self, it still leaves us with questions about how this illusion arises and what role it plays in our lives.
Personal pronouns are central to the human self system. In particular, it is the self-referential function of the first person pronoun that allows each of us to assert possession of brains, breath, beauty, buttons, brothers, etc. in a uniform way. To understand brains, breath, beauty, buttons, and brothers, one needs to understand anatomy and physiology, aesthetics, tailoring, and psychology. To understand how one asserts a claim on each of them one need only understand language. Thus I'm not concerned with understanding the "brother" component of the phrase "my brother." I'm only interested in the "my" component, which functions the same in that phrase as it does in similar phrases such as "my body," "my brain," or, for that matter, "my button." If we can understand the mechanism of self reference we are well on the way to explaining the illusion of a single Self at the center of our activity and Being.
Consequently I devote the single largest section of this essay to a detailed account of how a nervous system implements personal pronouns. However, I begin by laying some foundations and then move to discussions of the neural self and the problem of multiple personality disorder. We follow with the question facial imitation in neonates and some experiments Piaget conducted on children's awareness of their body movements. At this point we are ready for the pronoun discussion, after which we revisit Piaget's experiments and then move on to consider Vygotsky's account of language acquisition as involving the internalization of an other. The penultimate section is about how we animate the imaginary characters in our fictions. We conclude with an exorcism.
It goes without saying that this essay is speculative. The explanations I offer are the best I am able to construct given the materials available to me. While I hope they turn out to be true in the large, if not in detail, that possible truth is not the point of this exercise. What is important is that the explanations be interesting and significant, that they have the capacity to point us toward the truth. They can do that even if they are not themselves true. After all, Columbus discovered a new world for the peoples of Renaissance Europe despite the fact that he had only intended to establish a new route to a distant part of the known world. If attempts to falsify or extend these ideas end up replacing them with better ideas, they will have served their purpose.
2. Self and Other in the WorldCaveat: The detailed discussion of personal pronouns in sections 6.1, 6.2, and 6.3 is technical cognitive science and so may seem strange to some PSYART readers. You may want to skim lightly through those sections to get a sense of the model and then return to the discussion, with full attention, starting at section 6.4.
The brain is a part of the body, guiding it as it makes its way in the world. Figure 1 represents this relationship in the simplest possible way. Given this diagram it would be natural, then, to imagine the self as a box within the brain. We need not imagine that this neural self is some single continuous neural structure; it may well be many neural structures widely scattered about the brain. But for the simple purposes of our diagram, we could easily represent this complex set of neural structures as a single box.
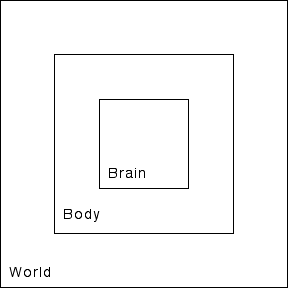
Figure 1: World, Body, Brain
That diagram would be a useful way of conceptualizing the basic physical relationships between the World, the Body, the Brain, and the Self, but it is not a useful way of thinking about the functional relationships. This requires a rather different diagram:

Figure 2: CNS in Two Environments
The point of this diagram is simple: The central nervous system (CNS), consisting of the brain and the spinal cord, functions in two domains external to it, the internal milieu and the external world. The CNS monitors the external world through various sensory systems--vision, sound, smell, touch, taste--and uses the skeletal muscle system to move the body and thereby act in and on that external world. The CNS also monitor's the body's internal milieu (including tracking concentration of various chemicals in the body) and affects that milieu through its control over the endocrine (e.g. adrenal) and exocrine (e.g. digestive) glands and the smooth muscles of the gut, blood vessels, and heart. (The shaded portion of the box representing the CNS represents the CNS systems involved in regulating the internal milieu.)
Physically, the CNS and the internal milieu are components of the same physical system, the body. In contrast to Figure 1, this diagram deliberately suppresses this obvious connection to emphasize the functional point, that the internal milieu is as external to the CNS as is the external world. Of course, the organism's survival, and hence the CNS's survival, depends on the integrity of that internal milieu. The CNS regulates the relationship between the external world and the interior milieu on behalf of that milieu.
If we examine the evolution of the nervous system we see that by far the greatest growth and elaboration has been in the structures and systems for observing and acting in the external world. The interior milieu of a human being is pretty much the same as the interior milieu of a rat, but their capacities for perception and action in the external world are quite different. Consequently, most of the differences between a rat's brain and a human's brain have to do with externally-directed activities, not with their internally directed functions. In the large, the same is true of humans and fish, though fish, being ectotherms, do not need the equipment for temperature regulation that all mammals require. Basically, as animals have evolved, the older neural structures are retained and carried forward, though sometimes they are put to new uses, and new ones added in as needed. Most of the new structures are externally oriented (cf. Bowsher 1973, Jerison 1976, Sarnat and Netsky 1974).
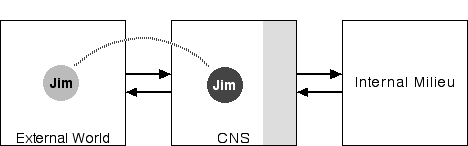
Figure 3: Jim in the World
Now consider Figure 3, which depicts a person, Jim, in the external world and the neural representation of that person--notice the dotted line indicating that relationship. While the neural representation of other people will be complex, our immediate purposes are such that all that complexity is adequately represented by a simple circle.
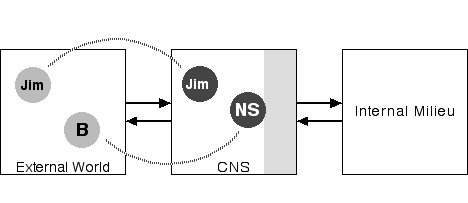
Figure 4: Self and Other in the World
With Figure 4, things become interesting. In this diagram B is the body of the person whose nervous system we are examining; the person who is contemplating Jim. What is their body doing in the external world on an even footing with Jim? The answer is really quite simple and straightforward. Imagine that you are looking at your leg. That leg is in the external world just as Jim is. You are examining that leg through your visual system just as you examine Jim. If you notice a cut in your leg you may touch it with your hand in the process of examining the cut. And when you put a bandage on your cut, you are using your motor system to act on your leg as though it were an external object, as Jim's leg most certainly is. You are your body, but the division of the nervous system directed toward the external world is quite capable of treating your body as an object on a footing with other objects. Correspondingly, the neural self, the NS (see Damasio 1994, 1999) in the CNS, is the neural representation of that body.
The obvious limitation of this diagram, of course, is that it fails adequately to depict the privileged relationship between oneself and one's nervous system, though I have indicated one aspect of that privilege by nudging the neural self against the shading representing CNS structures regulating the internal milieu. There is more to it than that, of course. You can move your muscles, but not someone else's. You can see only some parts of your body (unless you have a mirror) while all parts of other people's bodies are visible. And you can see through your eyes, hear through your ears, etc. but not with the eyes and ears of others. Thus the neural self is linked to the rest of the nervous system, and to the body, in a distinctly different way from representations of others. This difference is, of course, very important, and we will return to it a bit later in this essay. But, for the moment, I'm interested in the fact that, in a certain way, to a certain extent, our bodies exist in the external environment of the nervous system and so, in a limited sense, are on the same ground as other bodies.
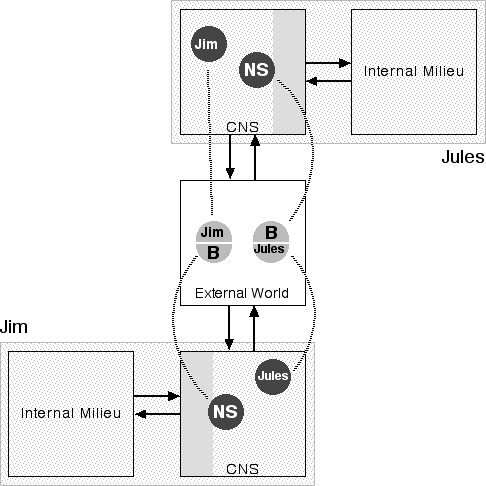
Figure 5: Jules and Jim
Imagine, for example (Figure 5), Jules and Jim involved in the kind of mutual grooming so beloved of chimpanzees. To belabor the obvious, each has his own body, CNS, and interior milieu. And each has the appropriate representations of his own body and of the other's. For this reason the two objects in the External World are given two labels as appropriate to how they are regarded by the two people. Thus the Jules object in the External World (at the middle of the diagram) is simply body (B) to Jules and is represented within Jules' CNS by the neural self. However, from Jim's point of view, Jules is a completely independent person and so appears as Jules and is represented within Jim's CNS as such. And so it goes with the Jim object as well.
As this grooming proceeds, one sees one's hands moving over the other's skin, out there--though not far--at the same place in the world. One's kinesthetic sense feels the motions of one's own hands and fingers while one's haptic sense feels the surface of the other's skin. One regulates one's actions through the intricate interplay of one's body and the other's while the other is doing the same. The object, of course, of this particular reciprocal interplay is to bring about a consonance between your interior milieu and theirs. This activity would be impossible if one's CNS did not, to some extent, represent one's own body in the same terms that it represents the bodies of others. To interact in a common space, that common space must be perceived and one must be perceived in it on the same footing as others.
Now, imagine that you are at a performance of Shakespeare's Antony and Cleopatra. You see actors on the stage playing the parts of Antony and Cleopatra. Figure 6 depicts this situation from the point of view of a playgoer:
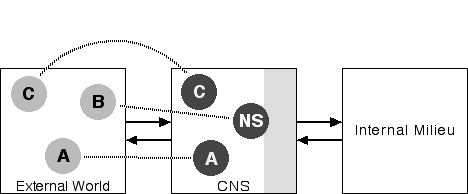
Figure 6: Play Acting
This diagram is the same as Figure 4 except that Jim, and his representation in your CNS, has been replaced by the characters of Antony and Cleopatra and their representations in your CNS. To the extent that you are not absorbed in experiencing the play you are aware of your body, in its seat, in the theater. But if the performance is a good one, and you are in a receptive mood, your attention will be taken up with Antony and Cleopatra on the stage. And your interior milieu will be responding to their actions. Your real body will have become a virtual body enacting, by turns, Antony's and Cleopatra's fictional bodies.
We will return to this bit of magic near the end of this essay. For now, it is time to think a bit more about the neural self.
3. The Neural Self : Damasio, Kilmer-McCulloch, and Dissociated Identity Disorder
In two recent books Antonio Damasio (1994, 1999b) has articulated a theory of the neural self. Damasio distinguishes between a core facet that is an integrated representation of one's body states and an autobiographical facet (1994, 236ff.; 1999, 171ff.). These selves--Damasio does refer to these systems as selves even as he refers to the neural self to mean both of these systems--are best conceived as processes, not things, and are subserved by extensive networks of interlinked neural areas. They are not physically continuous neural modules; the simple NS circle of the previous diagrams is thus a gross simplification (nor did the previous discussion say anything about the autobiographical self). Neither of these processes is the master process that runs the whole show--Damasio rejects the notion of such a process. As its name suggests, the autobiographical self organizes the historical events of one's life and imagines future events. The core self organizes sensations from the body's interior milieu and somesthetic and kinesthetic senses into an on-going evaluation of one's current body state.
Damasio's model is a rich one, but the purposes of this essay are most directly served by considering a single facet of it by considering anosognosia, a condition which disrupts the core self in a striking way (Damasio 1994, 62ff., 1999b, 209ff.). In this condition patients suffer brain damage that leaves them partially paralyzed, but they are completely unaware of their deficit and will deny that anything is wrong with them. Such, of course, is not always the case. It is possible, and common, to suffer brain damage that leaves one partially paralyzed but that doesn't spare one's awareness of that paralysis. It is this lost awareness of one's deficit that is characteristic of anosognosia.
Anosognosia generally occurs with extensive damage to the right hemisphere, leaving the left half of one's body extensively paralyzed--you'll recall that each cerebral hemisphere accepts input from the contralateral side of the body and sends output to that same side. The same pattern of damage to the left hemisphere will result in similar paralysis to the right half of the body but there is no lost awareness of one's disability. Anosognosia is a disruption of the integrated sense of one's body that is the foundation of the core neural self. What makes anosognosia so interesting to me is that it suggests a distinction primary neural systems that "run" the body and secondary systems that monitor the activities of those primary systems--a distinction we'll explore a bit later in conjunction with some experiments by Jean Piaget.
Anosognosia is a condition affecting the topology of the neural self, what is represented and what neural structures subserve that representation. There is another aspect of the neural self, one that has to do with the continuity and coherence of the representation. We can approach this issue by considering dissociative identity disorder (DID), an extreme pathology in which the neural self is fractured. In DID, also known as multiple personality disorder, one biological individual exhibits several different identities, each having different memories and personal style. In Thigpen and Cleckley's (1957) classic study Eve had three personalities; Schreiber's (1973) Sybil had sixteen (see also Rappaport 1971, Stoller 1973). Although there has been some controversy over whether or not DID is real or simply the effect of zealous therapeutic invention and intervention, there is no doubt that at least some cases are genuine (Schachter 1996, 236-242, Spiegel 1995, 135-138).
These different identities have different personal histories. The events in one personal history typically are unknown to the other histories. Each identity will have blank periods in its history, intervals, obviously, where another identity was being enacted. And the different "persons" are often unaware of one another. Further, the different identities seem to have different personal styles, different modes of speech, of movement, of dress, and so forth. Thus both the core and autobiographical selves seem to be riven. Using the conventions we employed above, Figure 7 is a simple depiction of DID:
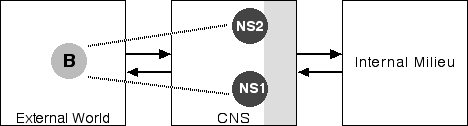
Figure 7: Dissociative Identity Disorder
Notice that we now have two neural selves, NS1 and NS2, corresponding to two different identities. Of course, these two selves exist in the same body, so we have only one corresponding body in the external world.
We do not, so far as I know, understand why or how DID happens. It is not, however, the result of the sort of gross destruction of brain tissue that underlies anosognosia. One might imagine, for example, that the different selves reside in distinctly different patches of neural tissue, a speculation that Damasio (1999b, 355) himself has suggested for the autobiographical self (though he presents no evidence). This suggestion, however, has at least one problem: How does the nervous system switch from one identity to another? There is another way of explaining DID, equally speculative and equally without specific evidence, that eliminates this particular problem.
Noting that different identities seem to favor different moods and that "memories established in one mood state are often more readily recalled in that same mood state than in a different one," Daniel Schachter (1996, 238) suggests that "different moods and roles come to be labeled with separate names" (cf. LeDoux 1996, 211-212, McGaugh 1995, Cahill and McGaugh 1996). Different selves emerge to handle different desires and emotions. This suggests problems with brain neurochemistry, for our moods, emotions, and desires are subserved by complex chemical interactions in the nervous system (Bremmer et al. 1996, Freeman 1995, 117ff.; Joseph 1999, Panksepp 1998, 1999). The distribution of many of these neurochemicals is regulated by brain stem nuclei, at least some of which are in the system which Damasio (1999a, 1999b) calls the proto-self--which also includes cortical structures. While the activities of the core and autobiographical selves are conscious, those of the proto-self are not (though others have a different view of the structures of this proto-self, e.g. Panksepp 1998, 309ff.). However, the structures of the proto-self help constitute the conscious states in which the core and autobiographical selves operate (cf. Hobson 1999a, 1999b).
Among these brain stem nuclei are those of the reticular formation (RF), one of the oldest structures in the brain. Classically (Moruzzi and Magoun 1949) the reticular formation has been associated with sleep and arousal. However, others have argued that the RF plays a broader role. Thus Vanderwolf and Robinson (1981) have argued that the RF exerts a general role in the control of adaptive behavior through its ability to influence the cortex. More recently, Damasio has argued that the RF and closely associated structures play a critical role in "managing body states and representing current body states. Those activities are not incidental to the brain stem's well-established activation role: they may be the reason why such an activation role has been maintained evolutionarily and why it is primarily operated from that region" (Damasio 1999, 274).
These views are reminiscent of a very elegant model proposed by William Kilmer and Warren McCulloch, one of the "first models of decision making in neural circuitry to explicitly opt for cooperative computation, rather than executive control" (Amari and Arbib 1977, 119). Noting that "No animal can, for instance, fight, go to sleep, run away, and make love all at once." Kilmer-McCulloch went on to list fifteen "mutually incompatible modes of vertebrate behavior," all of them as basic as those already mentioned (Kilmer, McCulloch, and Blum 1969, 279). The exact number and identity of these modes is not important. What is important is that, at all times, an animal must be in one of these modes, and only one of them. Kilmer-McCulloch hypothesized that it was the RF that determined which mode the animal was committed to at any moment (cf. Benzon and Hays 1988, 296-298). The RF has extensive afferent connections from the rest of the brain and the structure of its internal connections seems well-suited to making a global evaluation of those inputs, thereby assessing the current state of the organism. The RF also has extensive efferent connections to the rest of the brain and is thus in a position to affect its state (cf. Panksepp 1999, 21ff.; Green 1999, 43). Specifically, the RF is in a position to affect the neurochemical ambiance of cortical tissue and thus can affect just which synapses are most arousable at any given moment.
In this view, if the animal is in eating mode, the world becomes an array of objects and events that either promise something to eat, or interfere with that promise. The cortical patterns most relevant to eating are readily arousable while those less relevant are less arousable. Similarly, if the animal is in mating mode, then the world becomes a set of opportunities for sexual satisfaction or frustration. The RF commits the brain to a pattern of activation that is suitable to the mode and the animal then seeks its satisfaction. Just what it does is not directly determined by the RF; that depends on other neural centers. Figure 8 is a highly schematic representation of mode:
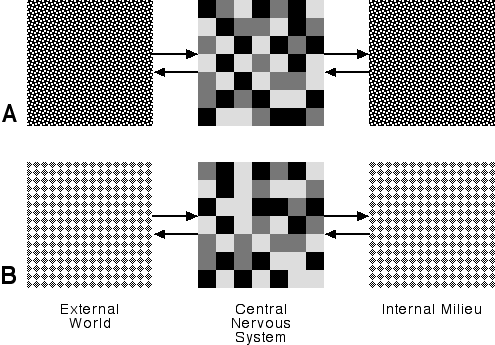
Figure 8: Behavioral Mode
Here we see two different modes, A and B. I've used some arbitrary pattern to color the internal milieu and external world for each mode, thus indicating that the animal is committed to bringing about a certain kind of consonance between its inner milieu and the external world. I've depicted the modal arousal of the CNS by varying patterns of shaded squares, where the shading corresponds to level of arousal. I've used three levels of arousal, but there could be only two, there could be seven, or level of arousal could be continuously variable. For our immediate purposes it makes no difference. What's important is that we have two distinctly different patterns of arousal corresponding to two behavioral modes.
My suggestion about DID, then, is that the mechanism that switches between one identity and another is fundamentally neurochemical. Each identity favors a particular mode, or, more likely, a set of modes. An identity becomes regnant when brain neurochemistry favors it. The perceptions and memories relevant to the modes of that identity will become easily arousable while those relevant to other modes will be all but impossible to arouse. Among individuals unaffected by DID the neurochemical milieu will bias cortical tissue toward a particular set of perceptions and memories but will not necessarily make other perceptions and memories impossible to reconstruct. In the case of DID this neurochemical process is taken to an extreme where whole ranges of perceptions and memories become absolutely unavailable depending on what neurochemicals are currently active. The state space of the brain has become fractured along neurochemical lines, breaking the self into many selves.1
Note that this explanation of DID not only tells us how the brain switches from one identity to another--RF control over cortical arousal--but also suggests that the various neural selves do not have be in physically separate tissue. They can exist within the same volume of neural tissue for they are differentiated by chemical sensitivity, not by location.
Of course, one doesn't have to think about this model too long before suspecting that it gives us more than we've bargained for. After all, neurochemistry is known to be implicated in various neurological and psychiatric problems, and one can easily imagine it to be implicated in problems where we currently have no specific knowledge. Part of the way out of this difficulty is simply to point to the complexity of neurochemistry. There are well over 100 known neuroactive chemicals (Hobson 1999a, 159). That leaves plenty of room for a wide range of pathological conditions, only one or a few of which will produce DID. The trick, of course, is to identify just which disruption is responsible for DID. That job is surely beyond the scope of this essay. (However, later in the essay we will return to DID to refine our speculation just a bit.)
The more general point, then, is however we theorize about the neural self, we must consider more than topological issues. It is not enough to know what and where in the brain the body state and autobiography are represented. One does not automatically have access to all the events the brain has registered. Autobiographical continuity is not given in the nature of the nervous system. The continuity and coherence of the neural self depends on complex matters of the neurochemistry of mood and emotion.
I would like to conclude this section by counterpointing DID against the perfectly normal activity of playacting (recall Figure 6). In playacting one deliberately assumes a different identity. One chooses to act like another and does so for a limited period of time. When the time is over one returns to one's own identity without, generally, ever having completely lost touch with that identity. Playacting is thus different from identity switching in DID, which is involuntary.
Children routinely engage in pretense during play. While adults are less likely to engage in playacting, one can certainly argue that reading novels and stories and seeing movies and plays involves something very like pretending to be someone else. You may not enact another person through gesture and voice, but you identify with fictional characters, enacting their feelings and desires in your nervous system, in your core self, and constructing autobiographies for them. Just as you can, in appropriate circumstances, reconstruct events from your past so vividly that you reexperience the feelings that attended them, so you can treat events in the life of a fictional character as though that life were your own and thereby experience that imaginary life as your own. Beyond this, of course, skilled actors can enter into a role quite deeply, creating physical and vocal styles appropriate to the character, imagining a lifetime of events in the character's life, not just those depicted in the script. In the case of a good actor, the transformation can be so great that one is confronted with the question of whether or not the actor becomes the character.
I have more to say about playacting but I want to say it after we have had a chance to delve into the cognitive mechanisms surrounding the personal pronouns. Those mechanisms will tell us a bit how a nervous system can pretend to be someone else. That investigation, for the most part, is about operations at the interface between the proto-self and the core self. With that discussion in hand we can revisit these neurochemical matters, concluding with the speculation that ritual and art contribute to the maintenance of an internal milieu which is not neurochemically fractured.
For now, it is time to consider the infant and how she interacts with others even in the first hour of life. That is where we will find the mechanisms upon which we will construct our account of the personal pronoun system.
* * * *
A Note on Conceptual Frameworks: David Hays and I have developed a fairly extensive cognitive model organized on several levels, which we choose to call degrees (Hays 1981, Benzon 1978, Benzon and Hays 1988). Henceforth, when I use the term "degree" I will be referring to that model. I use this model as the most general framework for this essay and, in particular, for the technical detail of the model for first and second person pronouns. Damasio's theory of the neural self, of course, is the other major framework for this discussion. Given that Damasio's model has been developed for rather different purposes, and employs a different evidentiary base, there is no clean correspondence between his model and ours. Roughly speaking, our modal degree contains at least some of the subcortical components of Damasio's proto-self. Our sensorimotor degree would handle some of the functions of both the proto-self and the core-self, but none of the functions of the autobiographical self. His autobiographical self would be implemented in our episodic and gnomonic degrees, though I suspect the core self has a gnomonic aspect as well. (See Note 9 for further remarks on the episodic and gnomonic degrees.)
The table below summarizes the approximate relationships between Damasio's theory and the Hays-Benzon model. In particular, the neural correlates more accurately reflect the Hays-Benzon model, which is about perception, action, and cognition in general, than Damasio's model. The notion of primary, secondary, and tertiary cortex is that of A. R. Luria (1973).
| Hays-Benzon | Neural | Damasio |
|---|---|---|
| modal | brainstem | proto-self |
| sensorimotor | limbic system basal ganglia cerebellum |
proto, core |
| systemic | primary, or projection cortex |
proto, core |
| episodic | secondary cortex (temporal, frontal) |
autobiographical |
| gnomonic | tertiary cortex (prefrontal, parietal) |
autobiographical |
* * * *
4. What the Toad's Brain Tells the Infant's Tongue
Consider the task of facial recognition. In the larger compass of this essay, facial recognition is important because it is at the core of social life. If you can't recognize people, you have little chance of interacting with them, and regulating interpersonal interaction is one of the Self's paramount responsibilities.
This situation is essentially the one we had diagrammed in Figure 5. Then we were imagining two adults interacting, now we are imagining an adult and an infant. The adult's nervous system is fully mature while the infant's is not. The facial recognition task is simple; the infant must recognize the adult's face as a face and act and interact accordingly. Once we've reviewed a particular model for facial recognition we will speculate about facial imitation.
Mark Johnson (the neuroscientist, not the philosopher) offers a developmental model for facial recognition in which a more sophisticated system emerges from a prior and simpler system. Johnson hypothesizes that facial recognition develops in two stages (Johnson 1997, 113):
. . . the first process consists of a system accessed via the subcortical visuo-motor pathway (but possibly also involving some of the deeper, earlier developing, cortical layers) that is responsible for the preferential tracking of faces in newborns. However, the influence of this system over behavior declines (possibly due to inhibition by developing cortical circuits) during the second month of life. The second brain system depends upon cortical maturity, and exposure to faces over the first month or two, and begins to control infant orienting preferences from around 2 or 3 months of age.
As the subcortical system is operative at birth it must have been "programmed" by the genes. Its operations are of the modal degree in the Hays-Benzon model. The second system is not so programmed; its operations are of the sensorimotor degree. The subcortical system primes the behavioral pump and guides the infant in actively exploring the environment. In the course of this exploration the sensorimotor (cortical) system begins maturing and provides the infant with a more finely differentiated capacity for facial recognition and discrimination than was possible to the modal degree alone. The capabilities of the modal degree are limited to what the genome can specify without any "knowledge" of the external world beyond what it acquired through the indirect route of biological evolution.
However important it is, facial recognition and discrimination alone do not create a social life for the infant. The infant must be able to do something to hold up her side of the interaction. One of the things infants do is to imitate others, which those others often find to be irresistibly fetching.
Lore has it that imitation is very important in primate life--monkey see, monkey do. There is a great deal of research which supports this view (Hinde 1974; Tomasello and Call 1997). Imitation is also important in human life (e.g. Piaget 1962). Through imitation we learn new skills and we also learn how others negotiate their way in the world. The regulation of imitative activity is surely under the purview of the Self system. I find the logic of Johnson's account of facial recognition so very attractive that I would like it to apply to imitation as well. We know, in fact, that human infants can reliably imitate adult facial expressions in the first days of life, even as young as 42 minutes (Meltzoff and Moore 1995, Bower 1977). For example, if you look at a neonate and stick your tongue out, the infant will imitate your action. The same for eye fluttering, lip protrusion, mouth opening, and finger movement. This happens so early in life that it seems unlikely that infants have learned how to do this. Rather, such behavior must be innate.
One might well wonder just how the genes could accomplish this bit of programming. I certainly don't have an answer to that question, but I can make a suggestion which is grounded in the fact that the human genome is heir to millions upon millions of years of vertebrate evolution. I suggest that the information processing demands of this simple kind of imitation seem to be on a par with the information processing activities of a toad zapping flies out of the air with its tongue (Ewert 1974). Working in the tradition of Warren McCulloch's classic account of "What the Frog's Eye Tells the Frog's Brain" (Lettvin et al. 1959), Jörg-Peter Ewert studied the visual mechanisms through which a toad distinguishes food from foe and responds accordingly. The visual discrimination is based on simple point and edge configurations, direction of stimulus movement, and contrast between foreground and background. A small horizontal object (i.e. worm-like) moving horizontally is food, to which the toad responds by turning toward it and snapping at it with its tongue. Larger objects and vertically oriented objects signify danger, and caused the toad to crouch in avoidance.
It is not too difficult to imagine similar machinery in a human infant being able to detect a pair of eyes or a tongue. An eye is a small colored spot--the pupil and surrounding iris--against a white background, while a tongue is a horizontal pinkish object moving around beneath the eyes. Given that such sights are freely available in the infant's world, and there are generally no similar sights that are dangerous, it wouldn't require sophisticated visual processes to identify a person wagging her tongue directly at the infant. I should think that a toad-class visual system would be adequate to the task. And the motor programming which a toad uses to catch worms and flies would seem to be, if anything, more sophisticated than that which allows an infant to protrude her tongue in imitation of an adult's tongue protrusion--after all, the toad has to hit small moving objects. I am thus suggesting that the human infant's imitative competence is implemented through perceptual and motor patterns which have been inherited from our reptilian and amphibian ancestors and put to rather different use.
In making this suggestion I am invoking the logic of an argument Nicholas Humphrey made in his provocative account of "What the Frog's Eye Tells the Monkey's Brain" (1970). Humphrey did a series of experiments in which he destroyed the cortical visual areas of two monkeys. While they lost the ability to identify objects, they still had crude visual abilities. Humphrey concludes (p. 336):
Is there any sense in the idea that after removal of the visual cortex the monkey sees in some ways like a frog, as if the lesion produced a sort of phylogenetic regression? . . . It seems possible that in the orienting movements a toad makes to a fly we are indeed witnessing the primitive homologue of the visual-grasp eye-movements with which advanced mammals fixate visual targets and which are the only token of visually guided behavior left in the de-striate monkey. Although the subcortical visual structures of the monkey may have a significance for vision beyond what they can be supposed to have in an amphibian lacking a visual cortex, the operations of isolating a visual figure for the attention of the cortex and detecting a prey object for the attention of the tongue are formally the same.
And so it goes with basic facial recognition and imitation. The subcortical modal degree which underlies the neonate's limited capacities for social interaction has capabilities which are comparable to the midbrain systems of frog and toad. It is perhaps not so difficult to believe that the genome could retain and reconstruct those capacities though millions of years of evolution. Those neural structures and capacities of the Inner Toad are among the unmoved movers of human social interaction and are, I propose in a later section of this essay, critical to the development of the self structure. These mechanisms belong in Damasio's proto-self. They are critical to the development and function of the core and autobiographical selves, but they don't afford the fine-grained control of perception and behavior these systems have. As we will see a bit later, these mechanisms also form a critical portion of the larger mechanism implementing the first and second person pronouns.
5. Talk That Walk 1
So far we have discussed two degrees of behavioral organization. One degree is innate and appears to be implemented in subcortical systems. The second degree develops in the first year of life and appears to extend into the cortex. Now we turn our attention explicitly to higher degrees. These systems, presumably, would all be under neocortical control as well, though they are likely to have specific subcortical components.
Let us begin with an experiment conducted by Jean Piaget as part of an investigation into consciousness. In this experiment, children were asked to crawl for about 10 meters and then to describe what they had just done (Piaget 1976, 1ff.). Four-year olds generally said either that they first moved one arm, then the other, then one leg, then the other, or legs first and then arms. Piaget called this a Z pattern. That is not, in fact, how any of them actually crawled. What they actually did was either to first move one arm, then the opposite leg, then the other arm, then the opposite leg, or the same pattern beginning with a leg. Piaget called this an X pattern. It isn't until children are seven or older that they can describe this X pattern. The four year olds, with their linguistic capability, exhibit a third degree (which Hays and I call systemic) of behavioral organization beyond the two we have already discussed in the previous section while the older children, with the ability to describe the X pattern of physical movements, exhibit a fourth degree (which we call episodic).
What is striking is that the younger children's verbal account of such a basic act is simply wrong. In order to execute the crawl there must be some brain tissue devoted to schemas regulating the appropriate actions; for crawling isn't a spinal reflex. But those regulating schemas must in some way be distinct from the schemas underlying the younger children's verbal accounts, otherwise those accounts would be more accurate. My first point is simply that we are here dealing with two different neural schemas for the same action and that one of them is grossly simplified and thus incapable of actually regulating the behavior it represents. This is similar to the distinction we made when considering anosognosia, where both sets of schemas were destroyed, in contrast to other syndromes, which destroyed the motor system's regulating schemas (thus causing paralysis), but not the secondary schemas allowing knowledge of that paralysis. (Note that, while the term "schema" has been much used in psychology, I do not use it in any particular sense.)
Let us say that the neural representation actually regulating the walking is a sensorimotor schema while the neural representation underlying the verbal account is a cognitive schema of the systemic degree. In the Hays-Benzon model the cognitive degrees are derived from and grounded in sensorimotor perception and action, a conviction we share with cognitive linguists. The meaning of the word "crawl" thus is ultimately given by the sensorimotor schema for enacting a crawl, or recognizing it in another. Sensorimotor schemas are thus Gestalt or analog in nature; these are the image schemata beloved by cognitive linguists (Johnson 1987, Lakoff 1987, Lakoff and Johnson 1999). By contrast, cognitive schemas are propositional or digital in nature, corresponding to the concepts (Johnson 1987, 153-155) and propositions (Lakoff 1987, Johnson 1987) of the cognitive linguists. The verbal account is proximately driven by cognitive propositions, which do change over time so that the older child can call on richer propositional capacities than the younger child. However, no matter how much it changes, the cognitive representation cannot serve the function of actually regulating the physical action; it doesn't have the requisite detail nor the proper peripheral connections. Propositions aren't as rich as Gestalts, but they are often more parsimonious (see the discussion of finitization in Benzon and Hays 1988, 304ff.) and allow for more compact storage.
The sensorimotor schema for, e.g. crawling, is multimodal and thus distributed over several brain regions. It is motoric (sending impulses to muscle fibers), kinesthetic (detecting movement at the joints), haptic (sensing the surface of the ground). But it is also visual. For one thing, one senses the optic flow of the world as one moves through it (Gibson 1979). One can also see one's own limbs moving, and, perhaps more significantly, one can see others' limbs move as the other crawls (recall Figure 5). The cognitive schema is not specific to any of these modalities; it is responsive to each.
But this essay is not primarily about the difference between sensorimotor and cognitive representation. For now, all we need is the notion that we have two neural representations, one linked to action, the other to language. The same two types of representation would also exist for other actions such as walk, run, jump, grab, make a hook shot (a basketball maneuver), play a C# scale (on some musical instrument), and so forth. There are the sensorimotor patterns which execute the action, and recognize it in others; and there are the cognitive propositions which underlie verbal description.
Similarly, we can distinguish the between the cognitive propositions involving "see" and the neuropsychological process of seeing, of visually examining the world and identifying objects and events in it. It is a simple matter to say something like: "I see that Priscilla is wearing the new diamond and ruby pendant Elvis bought for her at Tiffany's." The actual seeing of that scene involves the interpretation of visual input in various regions of the cerebral cortex, scanning the eyes across a scene so as to bring various parts of it into central focus, and relating the scan patterns to the objects and events identified (cf. Benzon and Hays 1988, 304-308). We also recognize that others are "seeing" by attending to the direction of their gaze and to their eye movements (Baron-Cohen 1995): "Fred's looking at his dog Spot." I would imagine the cognitive "see" attaches equally well to the complex of processes by which we actually see or the perceptual schemas though which we recognize seeing in others; those are the neural representations which give meaning to the word. Talk of seeing is linked directly to the cognitive propositions. It is the propositions that would be directly linked to the sensorimotor schemas doing the actual seeing or identifying it in others.
So, the difference between actually crawling (sensorimotor) and talking of crawling (systemic) parallels the difference between actually seeing (sensorimotor) and talking of seeing (systemic). We could construct similar accounts for a range of verbs of sensation, perception, desire, mentation, and communication: touch, taste, hear, smell, talk, say, write, think, laugh, dream, imagine, and so forth. Each of these words designates something which is under central control by the brain, parcels of brain tissue interacting in order to get these things done. The cognitive patterns which underlie talk of such things are much simpler. To the extent that those cognitive patterns represent something which any person does, they can be used in representing one's own actions or other peoples' actions. Our autobiography will necessarily contain representations of others as well as of ourselves. Just as one can in effect, use the core self to "expand" the memory cues of one's own past actions into fully realized actions in the present, one could also use memory cues of others' past actions as the basis for one's present actions.
But how does one "switch" one's core self from enacting one's own life to imitating another's life? The answer to that question, I believe, turns on the cognitive underpinnings of the first and second person pronouns. One executes the switch by becoming "you" rather than "I". The next section is about the design of that switch.
6. First and Second Person Pronouns and the Self Structure
There is a standard sense of the first and second person pronouns--"I" and "you" in English--which simply treats them as shorthand for "the person who is speaking" and "the person being addressed" respectively. There is no doubt that pronouns are that, but such an account doesn't get to the heart of the matter. That is the conclusion John Lyons (1977, 636-646) reached in a thorough discussion of the grammatical category of person. In particular, he notes that "In so far as 'the speaker' and 'the hearer' are substitutable for 'I' and 'you' in ordinary English, they are conventionalized pseudo-descriptions which (like 'the author' and 'your lordship') depend for their interpretation upon our intuitive understanding of how person-deixis operates" (Lyons, 645). Citing Benveniste (1958/1971), he concludes that the grammatical category of person "introduces an ineradicable subjectivity into the semantic structure of natural languages" (Lyons, 646).
To get a better understanding of Lyons' point, let us consider an example. The following scrap of imagined dialog uses personal pronouns in the standard way:
How was your trip?
Fine. I saw Victoria Falls--you would have loved them--and visited my grandmother's old village.
Yes, I would have, though I'd have been afraid to get too close.
I was at first, but you get used to it.
Provided she is present at the scene of this conversation, a third party witness would not have to know the names of the two speakers in order to identify the referents of these utterances. The pronouns indicate those referents to anyone present at the conversation, but not to someone, for example, who only reads a written transcript without identifying introductions.
Now let's imagine the same dialog with the personal pronouns replaced by proper names.
How was Jack's trip?
Fine. Jack saw Victoria Falls--Jill would have loved them--and visited Jack's grandmother's old village.
Yes, Jill would have, though Jill would've been afraid to get too close.
Jack was at first, but a person gets used to it.
This version doesn't feel like people are in conversation with one another. Rather, it feels like people are making related statements in one another's presence but there is no sense that they are talking to one another. If the speakers are, in fact, Jack and Jill, then the various statements are self referential. It is, however, possible that the speakers are not Jack and Jill, but Fred and Ginger. In this case there is no self reference. However, nothing about the language itself tells you anything useful about the speech situation. The language is disembodied and profoundly asocial.
The effect is just as strange if you substitutes phrases such as "the speaker" and "the hearer."
How was the hearer's trip?
Fine. The speaker saw Victoria Falls--the hearer would have loved them--and visited the speaker's grandmother's old village.
Yes, the speaker would have, though the speaker have been afraid to get too close.
The speaker was at first, but a person gets used to it.
In this case the distancing we observed with proper names is compounded by the effort needed to change the reference of the substituted expressions as the identify of the speaker and hearer shifts from one conversational turn to another. And, as Lyons indicated, these expressions work only because we already know, intuitively, just what "speaker" and "hearer" mean. Without that prior intuitive understanding such dialog would be utterly hopeless.
The trick is to explicate that intuitive understanding by explicating the underlying mechanisms. Those mechanisms are both linguistic and social. They are at the interface between language, the core self, and the mechanisms of social interaction.
When you converse, not only must you attend to the words themselves but you must attend to the person you are talking with, picking up cues indicating whether or not they understand or approve and making appropriate adjustments. You need to know how and when to interrupt the flow of information to ask a question. You need to gauge how much of an answer to supply to questions. In short, you need to have a sense of the other's intentions in order to engage in conversation.
It is thus interesting to note that autistic individuals often have difficulty using personal pronouns properly, often reversing "I" and "you," as though "I" is a noun designating some specific other person while "you" is a noun designating the autistic individual (Bloom 2000, 79, 125). We now have a body of research indicating that autistic individuals have trouble attending to and following those perceptual cues which indicate a person's state of mind, such as direction of gaze (Baron-Cohen 1995). They seem incapable to treating others as subjects and thus their interpersonal interaction seems profoundly asocial.
The first and second person pronouns orient utterances in intersubjective space and thus allow speech to be a social act rather than a disembodied proclamation of propositions. To establish a provisional understanding of this process we need first to consider the nature of the linguistic sign, to understand it as a set of representations in neural tissue interacting through the processes of language.
6.1 The Linguistic Sign
First we need to establish a notation convention. The following diagram represents a neurofunctional schema network:
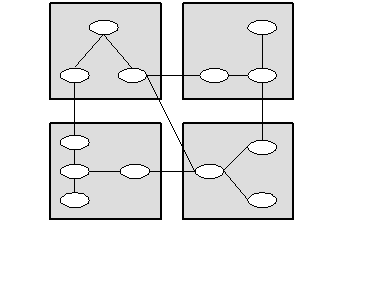
Figure 9: Neurofunctional Schema Network
The nodes (ellipses) represent schemas while the links connecting them represent relations between schemas. The rectangles represent neurofunctional areas (NFAs)--the various functionally differentiated cortical regions and subcortical nuclei recognized by the neurosciences.2Each schema is located in some NFA and each NFA will contain many schemas.
Thus conceived, a neurofunctional schema network is a kind of cognitive or semantic network, which is a well-known formalism for knowledge representation in cognitive science and artificial intelligence (Barr and Feigenbaum 1981, 180ff.). Much of the work David Hays and I have done on cognitive structures has made use of such networks (Hays 1976, 1981; Benzon 1976a, 1978). The concept of a neurofunctional cognitive network is a notation I have been considering in thinking about how cognitive networks might be implemented in neural networks, both those in human brains and the rather simplified devices implemented in digital computers. The general idea is that each node in a cognitive network represents some schema recognized by a neural net, where a collection of neural nets is a computational device that implements the cognitive network. Thus the nodes and links in an neurofunctional net should not be confused with the primitive neural elements that constitute a neural net. We should not, in general, expect there to be a transparent relationship between the elements (schemas) in a neurofunctional net and the elements (neurons) in the underlying neural net. We cannot say that just these elements in the neural net, and no others, implement this particular node in the cognitive net. Typically a given element in a neural net will contribute to many concepts or schemas recognized by the net and any particular concept will reflect output from a population of elements distributed throughout the local NFA.
The neural nets are organized into neurofunctional areas (NFAs) which have interconnections with one another (cf. Benzon and Hays 1988, 298-304, and the concept of constrained connectionism in Regier 1996). Thus any schema in a neurofunctional schema network exists in a particular NFA. A cognitive schema can be linked to any number of other cognitive schemas, either in the same or in different NFAs. The types of relationship possible between schemas in the same NFA seem to me quite distinct from the types of relationship possible between schemas in different NFAs. In particular, the activation of schemas in mutually interconnected NFAs will constrain one another so that only certain patterns of activation are possible. That is the force of this notational convention.
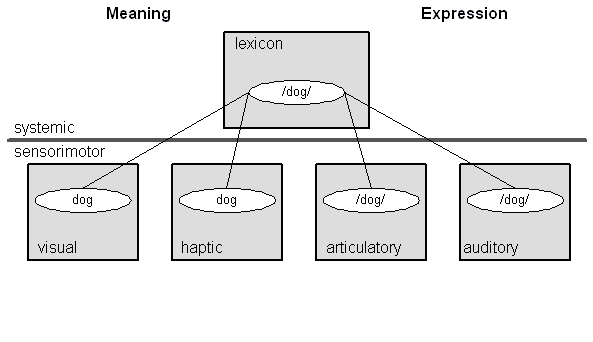
Figure 10: The Linguistic Sign
Now consider Figure 10, which shows the structure of the linguistic sign.3
At the systemic degree we have a lexical NFA; these are all the words of the language. At the sensorimotor degree we have general sensory NFAs for visual, haptic, olfactory, auditory, etc. schemas. These nodes contain schemas for the visual dog, the feel of its coat, the smell of its breath, and the sound of its bark. These give the word "dog" its meaning. In the language of cognitive linguistics, these nodes are the image schemas for dog (Johnson 1987, Lakoff and Johnson 1999). We also have sensorimotor NFAs for the articulatory gestures used to utter words and the auditory schemas used to comprehend them. Taken all together the schemas in these various NFAs constitute the linguistic sign (cf. Neisser 1976, 162-168). Our task, then, is to link the neural machinery of the linguistic sign to the neural machinery for managing the conversational interaction.
6. 2 Ego and Alter, I and You
Anthropologists and sociologists use the terms ego and alter when talking about social interaction. Ego designates the person from whose point of view the interaction is being considered while alter designates a person ego is interacting with. I propose that we use these terms as names for schemas which are part of our basic neural machinery for regulating social interaction. Consider Figure 11:
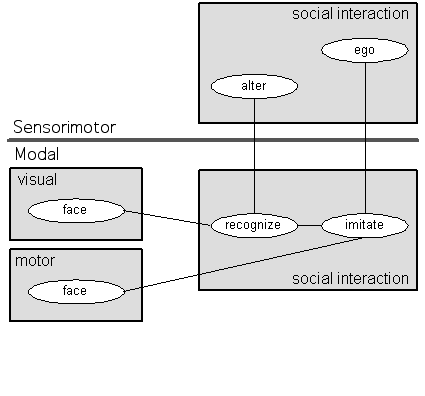
Figure 11: Ego and Alter
At the bottom we have the modal degree, the locus of innate systems for facial recognition and imitation. The face schema in the visual NFA is not, in fact, so simple as that node suggests. That schema is the innate program for facial recognition (cf. Fantz 1961). That schema would not be tuned to recognize a specific (parental) face, but, as a practical matter, human social organization is such that parents (and other close relatives) are the most likely bearers of the faces available for neonatal scrutiny. The point is that, initially, any face will do. That is what we want at the core of social life, a schema which can answer to anyone at all. Once the more sophisticated facial recognition circuits mature they would allow more differentiated treatment of others, but those innate schemas would still exist at the core of social interaction. Similarly, the face schema in the motor NFA is a program for manipulating one's own facial musculature. In the neonate, if this program is activated specifically in response to someone else's facial expression the facial gesture will be imitative. Given the prominence of the tongue in this behavior, this speculation has rich consequences for those contemplating a neuropsychological reconstruction of psychoanalytic thinking.
The social interaction NFA is similarly complex--considerably beyond what this simple diagram conveys--consisting of conditions and of actions to be taken when those conditions are met.4
The conditions are states (facial expressions, vocal gestures, touches, etc.) of the person one is interacting with and actions are the appropriate responses. In this particular case, on recognizing a face (condition), imitate it (action). The sensorimotor degree is above the modal degree and it too contains a social interaction NFA--perhaps in the cingulate cortex (Damasio 1999b). Among other things, it contains two nodes, ego and alter. These nodes are variables which are linked to the various innate interaction patterns in the subcortical social interaction NFA.
Alter is a role linked to the representation of the person with whom ego is interacting. Obviously, that person changes from one interaction to the next. The specific details about the person would be represented by nodes in other NFAs in the network. Similarly, the ego role is grounded in those circuits which orchestrate imitative response to other's facial presentations. In this sense, the ego node is but a particular limited function in the psyche, albeit an important one. Like its conceptual predecessors in this line of theorizing--the Self node (Benzon 1976a, 1976b) and the anchor (Benzon 1978, 177-79, 243-252, 315)--ego is not the whole of the Self; rather it is a point of reference in the interpretation and initiation of social signs and gestures. I should hasten to add that the ego node as defined in this theory is not equivalent to the psychoanalytic concept of the ego though, as an element of cognitive structure, it no doubt has a role to play in the wider mechanism of the psychoanalytic ego.5
The relatively circumscribed functional orbit of that ego node reflects an important aspect of this theoretical style and needs to be emphasized. As William Powers (1973, 1976) has pointed out, it is easy to draw circles and boxes and adorn them with impressive labels, like Self, or Interpersonal Affect Transducer, and to then think you have explained something. The ego node in Figure 11 is not such a construction. It should be quite obvious that these few diagrams are not complete plans for a brain, real or artificial. They specify some few necessary components and connections in such plans, and it is in that spirit that I offer them. These components exist at the interface between the proto-self (the modal components) and the core self (the sensorimotor and systemic components).
Now consider Figure 12, which is specifically concerned with first and second person pronouns, exemplified by "I" and "you":
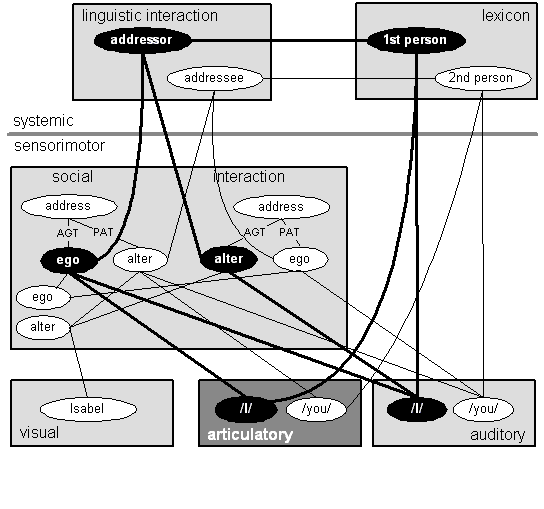
Figure 12: "I" and "you"
Notice that we now have the systemic and sensorimotor degrees; the modal degree still exists, of course, but I left if off the diagram to keep things a bit simpler. The ego and alter nodes at the lower left of the sensorimotor social interaction NFA are linked to conditions and actions in the modal social interaction NFA as indicated in Figure 11. Notice that alter is also linked to an Isabel schema in the sensorimotor visual NFA. Presumably our person is interacting with Isabel and this node is a schema--in fact, a complex of schemas--encoding Isabel's visual appearance.
The sensorimotor social interaction NFA in this diagram is the same one that was depicted in Figure 11; it may well be several different NFAs, but for the purposes of this argument we can think of them as one. That particular complexity is not our subject and so we may provisionally ignore it. What interests me are the ego and alter nodes and how they are linked to the addressor and addressee nodes in the linguistic interaction NFA, through them to the first person and second person nodes in the lexicon NFA and to the /I/ and /you/ nodes in the articulatory and auditory NFAs.
Notice that the social interaction NFA has two instances of the address program, a program for social vocalizing. In one instance ego is addressing alter, and, in the other, alter is addressing ego. AGT, for agent, and PAT, for patient, are case roles. The agent is the actor in some verb while the patient is an object of that action.6
If we think of the ego and alter nodes at the lower left as types, then the ego and alter nodes which are linked to the address programs are tokens of those types. First and second person pronouns are given their sense in patterns involving these tokens of ego and alter.
(Note that, in Figure 12 I have emphasized the first person nodes and links to make it easier to read the diagram. That emphasis has no formal significance. Similarly, I have made the articulatory NFA darker than the other NFAs to highlight the fact that it is linked to one instance of the address program, but not the other. That emphasis has no formal significance.)
The addressor node in the linguistic interaction NFA--which is, of course, part of the general social interaction system--is defined by the agent role in the address program regardless of whether ego or alter is the agent. Similarly, the addressee node is defined by the patient role. The addressor and addressee nodes are then realized in the lexicon by first person and second person respectively. The first person lexeme is realized by the auditory and articulatory schemas for /I/ while the second person lexeme is realized by the auditory and articulatory schemas for /you/. Those schemas are linked to the appropriate ego and alter nodes in the social interaction NFA.
In particular, notice that the auditory /I/ and /you/ are linked to ego and alter differently in the two instances of address and that the auditory /I/ and /you/ are linked only to the tokens of ego and alter for the address program where ego is speaking. When ego is speaking, that is, acting as the agent in the address program (left side of the social interaction NFA), the first person pronoun always indicates oneself while the second person pronoun always indicates the person one is addressing. Conversely, when ego is listening, that is, acting as the patient in the address program (right side of the social interaction NFA), the first person pronoun always designates the person who is addressing one while the second person pronoun always designates oneself. Thus the diagram shows the auditory /you/ being linked to ego and the auditory /I/ being linked to alter.
In a sense Figure 12 doesn't tell us anything we haven't known for a long time, that the reference of first and second pronoun shifts depending on who utters them. Indeed, they are often called shifters. What is new is the attempt to explain just how the auditory and articulatory schemas for these words are linked to mechanisms for social interaction. In particular, these mechanisms are associated with observations about neonatal social behavior and so they have some grounding in the innate equipment of the nervous system. These mechanisms are components in the linguistic mechanisms which produce the self-referential linguistic behavior we've observed all these years.
6.3 The Functional Center of the Self-Structure
Let us move this section toward closure by considering Figure 13, which represents a fragment of the mental machinery for one Isaac, who knows an Isabel. It repeats some things we've seen before and adds a few we haven't seen. Again, I have highlighted certain elements of the diagram so it is a little easier to pick out certain paths. I have highlighted the motor and articulatory NFAs to emphasize the fact that they are linked to ego, but not alter (compare with Jim and NS in Figure 4 where the motor and NFAs would be part of NS).
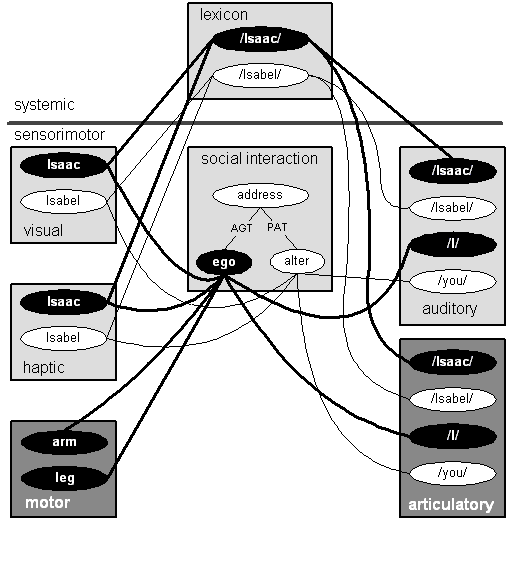
Figure 13: Functional Center of the Self-Structure
The motor NFA gives Isaac an arm and a leg with muscles he can flex and contract. We can imagine the visual and haptic schemas for Isaac and Isabel to be part of a rich network of information about them, physical appearance and manner, clothing, preferences of all sorts--food, movies, music, perhaps erotic technique, whatever, and episodes in personal history, which could be quite rich for both if they are long-term acquaintances, perhaps lovers, or spouses. The lexicon NFA displays nodes for /Isaac/ and /Isabel/, the names of our two imaginary subjects. These are linked to the appropriate visual, haptic, auditory, and articulatory NFAs.
The auditory and articulatory NFAs also depict /I/ and /you/. Notice that the ego node in the social interaction NFA is an agent in the address program (recall Figure 12); that is consistent with the links to the /I/ lexemes in the auditory and articulatory NFAs. The links between the visual and haptic Isaacs and ego, along with those between the motoric arm, leg and ego, and that between the articulatory /I/ and ego, indicate to us that this diagram depicts a highly idealized fragment of Isaac's brain rather than Isabel's. In the manner of Figure 12 we should imagine a token of ego in the patient role in another instance of the address program; that token of ego would be linked to /you/ lexemes. Ego would still be linked to Isaac in the visual and haptic NFAs and to the arm and leg in the motor NFA.
One thing needs to be emphasized. Discussions of the Self often talk of something called a self image, corresponding to what George Lakoff (1996) simply calls Self, or Self-of-I. This is, roughly speaking, linguistically accessible knowledge of oneself; the whole panoply of things William James (1890, 291ff.) had in mind when he spoke of a man's "clothes and his house, his wife and children, his ancestors and friends, his reputation and works, his lands and horses and yacht and bank-account." In this model that self image is carried in schemas in the visual, haptic, auditory, etc. NFAs which are linked to Isaac in the core self and, as well, the autobiographical self. The ego and alter nodes are not components of that self image. Rather, those nodes are components of the mental machinery through which the self image is brought to bear on social interaction and, as we will see in the next two sections, reflective thought. That puts them in what Lakoff calls the Subject, or Subject-of-I. In Damasio's terms, the Self-of-I would contain the autobiographical self and perhaps aspects of the core self as well. The ego and alter nodes would be components of the proto-self. They are unconscious components of the machinery that creates the conscious awareness of the core self and the autobiographical selves.
6.4 Banishing the Ghost
I would like to conclude this section by observing that we now have all the conceptual apparatus we need to argue that the notion of a unitary self is an illusion of language. More precisely, it is an illusion that arises when we reify self-referentiality. That reification becomes The Self.
The self-referential functions of the first person pronoun allows us to assert ownership over the entire inventory of things and events in one's life. One can thus imagine a very large collection of propositions about what "I want," what is "mine," things that "I did," and "my" stake in this or that endeavor. Stepping back from these propositions and examining them, it is easy to conclude that there must be some uniform and controlling agency behind this "I." This, of course, is a philosopher's job, for, as Lakoff and Johnson have pointed out, our common sense notions of the self allow for multiple agencies (Lakoff 1966, Lakoff and Johnson 1999, 267-289. Crudely put, our philosopher might reason like this:
Here we have the word "apple," and there we have an apple to which it refers.Similarly . . .
Here we have the word "I," and there we must find the Self to which it refers.
We have thus reified The Self and are off to the philosophical and psychological races, tracking down that Self which is the referent of the first person pronoun, looking for the essence that must give the word its meaning.
Note, however, that we have been able to conjure up a neural mechanism that works in such a way so as to produce those personal pronouns and their self-referential propositions. That is all we need in order to free ourselves from chasing after the elusive Self. Self-referential assertions find their meaning only in the process that produces and consumes them; that meaning is not some fixed essence, some Self. All we need to understand is how that process works. We know how "I" and "me" are produced. Beyond that, there is nothing more to understand.
It is one thing to describe the effects achieved by a mechanism. It is quite a different matter to explain how the mechanism achieves those effects. To describe the effects achieved by a Jacquard loom one need only examine the various fabrics and describe the patterns. It is quite a different matter to describe the loom's components and their interactions. It is the latter description we have been constructing for the pronoun system. Now that we know how the system works, we know all there is to know about self-reference.
7. Talk That Walk 2
Let us now revisit our child crawling and discoursing about that crawl. Figure 14 shows a bit of the network regulating Isabel's actions as she asserts that "I move my leg." Most of the NFAs and interconnections in this diagram are familiar from previous diagrams, the social interaction NFA containing the ego node, the articulatory NFA with its schemas for the vocal realization of words, the lexicon, and the linguistic interaction NFA containing the addressor node.
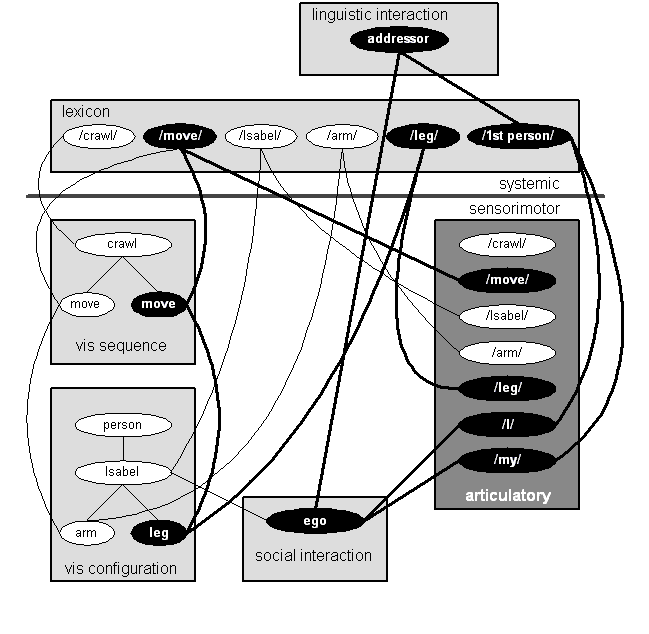
Figure 14: Isabel: "I move my leg."
The visual (vis) sequence and visual configuration NFAs would seem to be new. They are not. Rather, they are simply subregions of the sensorimotor visual NFA which has appeared in previous diagrams. The terms "configuration" and "sequence" are from William Powers (1973) and refer to specific levels in his servomechanical stack.7 Configuration schemas regulate simultaneous position in space, such as visual forms, or body postures. Sequence schemas regulate the succession of configurations in time.
The schema network in the visual configuration NFA can be translated roughly as follows: Isabel is an instance of person. Isabel has an arm and a leg. Of course, there are many other instances of person and most, if not all, of them have, not only an arm and a leg, but two of each, plus a plethora of other body parts. And these body parts are not just thrown in a pile in any old order. There is a definite structure which links them to one another. There is a large literature on such matters and so we can afford to leave those details aside for the purposes of this particular argument. The important point is that a given node in a schema network is linked to many other nodes directly and indirectly. The meaning of a node is a function of its position in the entire network; for that position determines how the node functions in perception, speech, thought, and action (cf. Lamb 1998).
The schema network in the visual sequence NFA asserts, roughly, that crawling involves moving an arm and a leg. The arm and leg being moved are represented in the visual configuration NFA and not directly present in the visual sequence NFA. Of course, there is more to crawling than just the movement of an arm and a leg; the other arm and leg must move as well, and the relative ordering of these motions must be represented. Those details are, like those of body parts and people, secondary to our present argument, though we should recall Piaget's experiments, which suggest we need at least two different accounts of crawl, one more accurate than the other.8
There is one more issue to deal with before getting to the point of this diagram. Why, if we are dealing with an utterance about one's own movement, are we deriving the content of that utterance from visual NFAs rather than motor NFAs? Here I am following Martin Sereno (1990) who suggests that the visual mode, which occupies 50% of the primate cortex, is the foundation of linguistic meaning. I am thus implicitly suggesting that talk about oneself is based on the language one learns for talking about others, language that assigns a privileged role to visual concepts. Beyond this, we might as well speculate that the difficulties that the three- and four-year-olds have in accurately describing crawling might be a consequence of deriving their description from visual schemas. In making a description the child simply does not pay attention to what her body does, but rather reports a poorly observed visual account of the action as she has observed others execute it. We can continue down this garden path of speculation by observing that the greater accuracy of the seven-year-old's account might derive from taking motor schemas into verbal account. That is, the greater sophistication afforded by the maturation of episodic degree structures permits, among other things, the explicit coordination of visual schemas of actions with the motor schemas which actually regulate those actions.
Whatever that case may be, let us turn our attention to the highlighted nodes and arcs in this diagram. This is the pattern of cognitive activity which underlies the assertion that "I move my leg." The content of the assertion derives from the social interaction NFA, containing the ego node, and from the two visual NFAs. The link between ego and the movement is not highlighted, but it is there in the diagram, which shows that ego is Isabel and the leg which is moving is her leg. That linkage is implicit in the cognitive structure, but plays no explicit role in formulating the sentence and so it is not highlighted. The lexemes which mediate between the semantic content and linguistic forms are at the top of the diagram in the lexicon NFA while the linguistic forms are in the articulatory NFA to the right; note the alternative realizations (/I/ and /my/) of the first person lexeme, which serve distinctly different syntactic functions. When you consider that we have yet to account for just how those linguistic forms get uttered in that particular order, it should be obvious that quite a bit of neural machinery is involved in producing this simple assertion.
What has become of the self structure? It is certainly there, but saying just what the Self is seems tricky. The self image in the two visual NFAs (and NFAs for other sensory channels as well) belongs to the core self and would also have links to the autobiographical self. The cognitive equipment which relates that image to the organism in which it is constructed seems to emanate from the ego node in the social interaction NFA.
That is a pretty weak formulation, "seems to emanate . . . " Yet it feels correct. We are dealing with a pattern of conceptual and perceptual relationships having a certain center. That center is not a (privileged) box in the mind; it is a point of orientation, no more and no less. The self structure, if it is anything, is an ordering of the world with respect to that point. And that ordering is not a static structure. It is dynamic and created from moment to moment. When we go to sleep that ordering dissipates. When we awaken, we must reestablish that ordering of the world and link the experiences of our wakening core self to scraps and shards of memory in our autobiographical self. When the connections have been made we know who we are and what we are about.
8. Says I to Myself . . .
Given this conception of self and language I now want to turn to Vygotsky's conception of thought as inner speech. The general idea is that as others direct the child's actions and perceptions through language, so the child learns to use language in controlling herself (Vygotsky 1962; Luria 1959). In effect, the child peoples her brain with an other and uses that other as a mechanism to control her own mind.
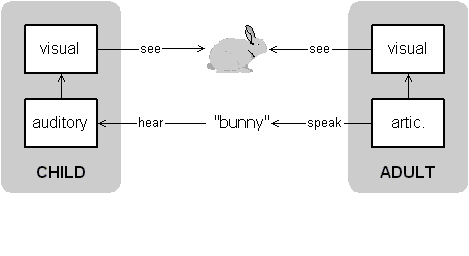
Figure 15: Adult directing child's attention to a bunny.
When a young child is requested to do something, the linguistic channel in the child's brain analyzes the acoustic input and activates the appropriate cognitive and perceptual schemas. The command, "Come here", will activate a plan for locomotion while the command, "Look at the bunny" (see Figure 15 for an informal representation), will activate a plan for seeing. The child knows that she is to execute the command because of the intonation pattern (Jakobson's conative function, 1960), which, presumably, is grounded in the various neural systems subserving social interaction. Upon receipt of that intonation pattern, the child's motor system is prepared to execute a pattern. As the content of the utterance is decoded the motor schema, whether for moving her body or looking in a certain direction, is executed. This sounds as though the infant is helpless in the face of intelligible commands from others, that she has no choice but to execute them. Initially, I believe, this is the case. The motor control center has no way of distinguishing between a command originating in the brain of another and a command originating within the child's own brain. Once the ability to make the distinction is learned, the word "no" enters the child's vocabulary as a means of marking autonomy (Church 1966, 101).
Not only can the child listen, she can also speak--though there is a lag between the child's capacity to understand language and the child's ability to produce it such that the child can understand more than she can talk about (Lenneberg 1967). If the child's utterance contains a command directed toward herself--and there is evidence on this (Vygotsky 1962; Luria 1959)--then she is using language to direct her activity in the way which others use language to direct her activity (see Figure 16). The route from acoustic analysis to the execution of the action is the same in both cases, only the utterance's point of origin is different. In one case the utterance originates with another, in the other case with the child herself.
Figure 16: Child talking to herself about a bunny.
The next developmental step, so Vygotsky's account goes, is that the child's self-directed speech becomes silent and internal (see Figure 17). In a word, it becomes what is ordinarily known as thinking (Benzon 1976a, cf. Lamb 1998, 181-182). Given that this process starts with language which others direct to the growing child and involves mental structures for coordinating language and social interaction, this would make thought, so understood, to be an inner dialog between virtual persons. It is thus not surprising that, in his investigation of the metaphor system governing folk conceptions of the self, George Lakoff (1996) found that we conceive the self to be a multiplicity of agents. And when we leave the world of folk psychology and enter the world of psychoanalytic thinking we find that the superego, the inner agent of social norms, develops as an internalization of parent figures (for example, see Freud 1960, 18-29; Erikson 1963, 256-57).
Figure 17: Child thinking about a bunny.
Let us then assume the existence of the mental machinery responsible for the process Vygotsky has outlined. This machinery operates on the self-structure as described in the account of personal pronouns. In the context of this more sophisticated machinery the ego and alter nodes are variables. Let us assume that this machinery creates what Gilles Fauconnier (1994) calls mental spaces; a mental space is an occasion for thinking, speaking, or writing.9
It is a temporary event; neurologically it would correspond to a particular pattern of brain activation.
A mental space can be real or hypothetical (Lakoff 1996, 95ff.). A real space is one representing real events, whether immediately present or recalled from the past. When activated in a real space the ego variable is set to equal the self-image while alter is set to equal the person with whom one is talking. This means that all the knowledge one has of that other is brought into play, whatever that means, through the operations of the alter variable. And, of course, all the knowledge one has of oneself is brought to bear through the ego variable. One constructs a hypothetical space to consider hypothetical situations about real people or, possibly, imaginary events about completely imaginary folks. When activated in a hypothetical space ego can be set to equal any person, as can alter. In particular, it is possible to set both ego and alter to the same value. When one is just thinking, ego and alter may both be set to the self-image, thus making such thinking into a conversation with oneself. When one is watching a performance of Antony and Cleopatra, ego may be set to Antony while alter is set to Cleopatra, or Caesar, or Enobarbus, whomever Antony is interacting with at the time--a matter we'll take up in the next section.
With this in mind, I would like to shift gears a bit and consider an image from my childhood. When I was seven or eight I was given a toy which we can call the COSMIC RAY GUN--I forget the real name. It had the shape of a pistol, but it didn't shoot toy bullets or even make toy noises. Rather, it was a device for projecting short film-loops on the wall. Each time you pulled the trigger it would advance the film one frame.
This toy came in a box and the box cover had some lettering which proclaimed the toy's name, COSMIC RAY GUN, a picture of the toy itself, and a picture of a boy holding a box. The boy's box had some (somewhat smaller) lettering, COSMIC RAY GUN, a (somewhat smaller) picture of the toy itself, and a (somewhat smaller) picture of a boy holding the box. This regress went as far as the resolution of the image would allow, but I was just clever enough to be able to imagine it going on and on and on without discernible end. That is to say, my brain was somehow able to imagine that each representation could itself become the object of a representation.
Let us give Isaac a COSMIC RAY GUN and imagine him holding the box and examining it. The image manages to embed an box within a box, etc. to the limit of the resolution of the printing technology. Thus seven-year-old Isaac can clearly see the first several terms in what you and I know to be a potentially infinite regression. The question before us is how it is that Isaac can get intimations of that infinity given that, at seven years, his capacity for abstract thought is quite limited. Isaac is looking at the box. In particular, he is looking at the smallest boy--call him Raygun Boy--in the picture.
Time 1) Set-up a real space in which Ego is set to Isaac and a hypothetical space in which alter is set to Raygun Boy. One is prepared, for example, to have a conversation with this imaginary companion.
Time 2) Now set ego to Raygun Boy. One is, in effect, prepared to speak on behalf of Raygun Boy. The system (i.e. Isaac's brain) now interprets the image before it as yet Another Raygun Boy on the cover of the COSMIC RAY GUN box, with its embedded series of images.
Time 3) Set alter to Isaac. Raygun Boy knows he is being observed by Isaac.
Time 4) Set ego to Isaac, who observes Raygun Boy on the cover of the toy box.
And so forth . . .
This process is not, in fact, a very coherent one. It begins to fall apart at step 3. The important point, however, is that by step 2 Isaac has managed to glimpse that there is, in principle, a series of boxes and boys beyond what he can actually see. Anything beyond that point is simply uncontrolled flailing about.
What is important, however, is that in this example we can begin to see a cognitive basis for a concept of an infinite recursive process, an abstract mathematical concept having nothing to do with social interaction. The concept of recursion is central to twentieth-century developments in mathematics, logic, computer science, and linguistics.10 That it should turn up in an investigation of the neuro-cognitive basis of the self is thus gratifying, though perhaps not terribly surprising. What is more significant is that that investigation did not presuppose recursion, but rather arrived at it though a particular construction.
8. Dissociative Identity Disorder, Again
We are now ready to revisit dissociative identity disorder. Given the concept of a neurofunctional schema net from section 6.1 we can visualize this as follows:
Figure 18: Neurochemical Bias
Figure 18 shows a fragment of network structure in some cortical neurofunctional area. In keeping with the notational convention, these nodes and arcs do not represent neurons, but rather patterns of activation distributed over many neurons. The arousability of nodes in the network depends on the neurochemical milieu of the cortical tissue realizing the network. Figure 18 depicts the pattern of arousability of the network under two different neurochemical regimes. I've indicated three levels of arousability but, as before (section 3, Figure 8) the number could be more. The difference between Figure 18A and 18B should be obvious. The nodes have Some nodes have the same arousability under both neurochemical regimes while others have different levels of arousability. Contrast this with Figure 19:
Figure 19: Neurochemical Fracture
Here fragments of the network simply disappear depending on neurochemical state. They are not arousable at all. This, I speculate, is the case with DID. Fragments of the neural self, both core and autobiographical, cease to be available depending on neurochemical state.
Given our model of the pronoun system we can further speculate about where, in terms of the model if not in specific neuroanatomical terms, this fracturing takes place. Schachter (1996, 238) had suggested that different moods have different names. That suggests that the lexical NFA of Figures 13 and 14 is one of the affected areas. Instead of one name one uses for oneself there are several, each specific to a particular neurochemical regime; this would, of course, affect other lexical items realized by the same tissue. I would also give attention to the linguistic interaction NFA of Figures 12 and 13. There would be several different addressor/addressee pairs, each neurochemically specific. Similarly, if the social interaction NFA of Figures 11, 12, 13, and was damaged, we would have neurochemically specific sets of ego/alter pairs.
In focusing on these particular NFAs I do not mean to imply that DID is a linguistic disorder. It is not. Note that the social interaction NFA of Figures 11, 12, 13, and 14 is not specifically linguistic at all. The NFAs we have been examining are critical to managing social interaction; they are at the interface between the proto-self, the core self, and language. Language, as the substrate of inner speech, is critical to mental life. Inner speech is the medium of silent biographical recall and contemplation, just as written language is the medium of diary and autobiography. If the critical structures at the junction of language and social interaction become neurochemically fractured, that could affect the much of the core and autobiographical components of the neural self.
9. Giving Life to Fictional Characters
How then, do real people give life to imaginary characters? To begin I want to consider a different question: How do we give current life to past experiences? To do this we must recall those experiences, and we must recall them vividly.
As far back at least as F. C. Bartlett's classic account of Remembering (1932), there is an intellectual tradition that sees remembering as active reconstruction rather than passive recall, as though one simply plays back a mental videotape. This tradition is quite alive in the contemporary cognitive and neurosciences (see papers in Shachter 1995). In particular, Damasio and others (1994, 96ff.; cf. Kosslyn and Konig 1995, 341-400, Posner and Raichle, 84-103) have noted that the many of brain areas involved in reconstructing the past are also those used in perception.11 Thus when we remember an event from our past we, in effect, recall a few cues, and then use them to reconstruct the event using some of the neural systems we used when we originally experienced the event.
Now, what is the difference between using one's diary as the basis for reconstructing past events and remembering those events without such an aid? Reading from the diary, of course, requires the use of the visual and language systems in ways not required by unaided recall, and the diary record may be more complete (but not necessarily so), but the neural process of reconstruction would be pretty much the same in both cases. The differences have to do with how one gets the cues on which to base the reconstruction, not with how one actually accomplishes that reconstruction.
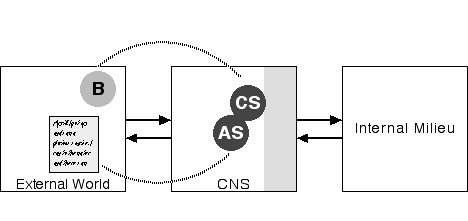
Figure 20: Dear Diary
Thus Figure 20 shows someone reading from their a where the diary is associated with that person's autobiographical self. The diary, of course, is an object in the external environment. We've now differentiated the neural self into its core and autobiographical subsystems. Now, what happens if, instead of one's own diary, one reads someone else's diary? What if one is reading a formal autobiography, biography, or history? The text depicted in the diagram changes, but the relationships do not. There may well be differences in the richness of the cues one readily summons to the words, but the process of reconstruction will be the same in all cases. In the manner discussed in the previous section, one sets ego equal to the person whose story one is reading, and one reads on, treating words about that person as though they came from one's own autobiographical self. As one constructs rich sensory events from those cues, one experiences the relevant portions of that reconstruction through one's own core self. That is what it means to identify with someone, to emphasize with them. You reconstruct the events of their life in your body and nervous system.
Linguistic accounts of other people's actions have much the same form as linguistic accounts of one's own actions. The verbs are the same as are the bodily participants governed by those verbs: arms and legs, eyes and ears, etc. One does not, of course, have direct access to another's sensorimotor system. One cannot directly see the world through another's eyes nor walk in it on their legs. But one can imitate them. Where the language says "walk" you execute the sensorimotor program for walking and the core self notes how that feels. Where the language says, "Look around and see," you execute the appropriate program, with the core self noting satisfaction, or not. And where the language says "he fell on his sword" you content yourself with a fake fall onto a blunted sword. The core self feels that as well and is no doubt grateful that the sword is blunt and that the fall was not a hard one.
From here, it is but a small step to identifying with imaginary people, with fictional characters. In all the cases we've considered already, the material one starts from is an account of something that someone experienced in real life. Now the initial material is not directly derived from real life. It is made-up. And the process by which one elaborates on the original material so as to produce a fully lived experience, that process does not depend in any way on the source of that original material. It can work just as well with imaginary materials. The fictional nature of literary characters may pose ontological, epistemological and moral problems for philosophers, but the problems they pose for psychologists seem little different from those posed by the presence of real people in journalistic, biographical, and historical texts. These are problems of realizing a full experience from various cues.
A recent PET study (Fink et al. 1996) conducted at London's Institute of Neurology is relevant to these issues. Subjects listened both to autobiographical statements and anecdotes and to similar material about a person they did not know. They were asked to imagine themselves in the described situation. In both autobiographical and non-autobiographical conditions there was considerable activation of the right and left temporal lobes in areas known to be associated with language (left) and episodic memory (right). However, the autobiograpical condition produced considerably more activation on the right, including limbic areas of the temporal lobe and the posterior cingulate area. These areas are known to be involved in emotion and social interaction, and the cingulate area plays a role in the core self (Damasio 1999b).
That autiobiographical material recruits a different and more extensive pattern of neural activation than non-autobiographical material is not surprising. After all, one's personal memories would be, in principle considerably richer than anecdotes about others. Yet, if our speculations above are to have merit, one would expect it possible for non-autobiographical material to recuit brain activation as extensive as that reported for biographhical material. The verbal stimuli in this case are certainly not very rich. The following two examples were given in the article:
When you were 15 you took part in a swimming marathon and succeeded to swim 10 miles.
He tore off his shirt to demonstrate his scars to the nurse.
Such statements are not comparable to a well-written biography or novel. Not only are events and incidents more extensively realized in such works, but we enter into and live with these works for a period of time during which we may read for an hour or more a day over a a week or more. In the case of a play or a movie we see fully realized characters in front of us and, of course, we are immersed in the experience for an hour or three.
Thus this study need not presage the death of the speculations we have been developing. On the contrary, it suggests that we have the experimental means at hand necessary to gather empirical evidence that bears on them. So, let us return to our speculations.
The process of identification and realization can reach an extraordinary depth when done by a skilled actor. Let us consider a passage from Constantin Stanislavski's An Actor Prepares. The book takes the form of an imaginary dialog between a distinguished director and actors taking lessons from him. At one point Stanislavski's alter ego suggests that "you are playing the scene in the last act of Hamlet where you throw yourself with your sword on your friend Paul here, who enacts the role of the King, and suddenly you are overwhelmed for the first time in your life with a lust for blood." Stanislavski (1948, 165) goes on to ask whether or not "it would be wise for an actor to give himself up to such spontaneous emotions as that" and to assert that:
. . . these direct, powerful and vivid emotions do not make their appearance on the stage in the way you think. They do not last over long periods or even for a single act. They flash out in short episodes, individual moments. In that form they are highly welcome. We can only hope that they will appear often, and help to sharpen the sincerity of our emotions, which is one of the most valuable elements in creative work. The unexpected quality of these spontaneous eruptions of feeling is an irresistible and moving force.
In such moments "playacting" has obviously driven the actor quite "deeply" into his nervous system, for such sparks of emotion necessarily require activity by subcortical neural mechanisms (see for example, Panksepp 1998, 1999), of the proto-self and the core self.
Along similar lines, consider these remarks that Leonard Bernstein (Epstein 1987, 52) made to a class of conducting students:
I don't know whether any of you have experienced that but it's what everyone in the world is always searching for. When it happens in conducting, it happens because you identify so completely with the composer, you've studied him so intently, that it's as though you've written the piece yourself. You completely forget who you are or where you are and you write the piece right there. You just make it up as though you never heard it before. Because you become that composer.
I always know when such a thing has happened because it takes me so long to come back. It takes four or five minutes to know what city I'm in, who the orchestra is, who are the people making all that noise behind me, who am I? It's a very great experience and it doesn't happen often enough.
What, in such a cases, is there to prevent a performer from becoming lost in the character, or composer? We could ask the same question of those religious celebrants who become possessed by a god or goddess and speak with their voice (for accounts of possession see Deren 1953, Friedson 1996, Rouget 1985). What is there to bring the celebrant back to her secular persona so she can do the mundane things a body needs to do in order to survive? What is there to prevent the actor or celebrant from succumbing to dissociative personality disorder?
Given the speculation about DID that we have developed above it does not seem likely that ritual or theatrical identification will result in pathological dissociation. As long as the critical areas of neural tissue have not become neurochemically fractured, the neural self will not splinter. However, some explanatory work remains.
Given that the neural self is not neurochemically fractured, we could invoke the ego/alter mechanisms we examined in our discussion of Isaac and Raygun Boy. Just as the performer can assume a role by setting ego to, e.g. Mark Antony, so the performer can return to himself by setting ego back to, e.g., Richard Burton. However, it isn't that simple. If it were, then it wouldn't have taken Bernstein several minutes to reestablish his current situation. For that matter, it takes time to enter into one's role as well. An actor doesn't just walk in off the street, step on stage, take a turn and simply become Mark Antony. Nor can a musician enter fully into performing persona in an instant. The transitions to and from one's performance persona take time. Whatever role ego/alter mechanism plays, one must also explain the minutes required to effect the transition between mundane self and performer. That such connections take minutes suggests that we're dealing with altering one's entire brain state, not just some few circuits between the proto-self and the core self.
Were I to look for an answer to this question I would look for it in the entire ritualized nature of the social process involved in intense rituals and performances. What allows the celebrants, performers, actors, and, for that matter, members of the audience, to come back to themselves is the fact that their interaction with one another is highly stylized and specially marked. They wear special ritual clothing, interact in a special ritual space, speak special ritual words--often in a special ritual language, and perform special ritual motions. Nothing is like its counterpart in the mundane world. Hence there is no possibility of being confused.
Experience is not ordinarily marked as Real or Imaginary in the way that television images are marked by a channel number and a logo periodically appearing on the screen. Ritual, by contrast, marks everything by making objects and actions visibly different from their mundane counterparts. The whole neural system is affected by this pervasive ritual marking. We are dealing with the states of the entire nervous systems of people in ritualized interaction with one another.
I would like to go beyond this to speculate that such performances play a role in helping to establish and maintain the subjective continuity of the neural self despite the neurochemical swings which, when out of control, result in such psychopathology as dissociative identity disorder (cf. Benzon 1981, 1993a, Hays 1992). By entering into a wide variety of emotional states (with their various neurochemical substrates) in a socially controlled situation, individuals in a community create an "equal access zone" in mental space where each can experience and contemplate extremes of joy and anger, tenderness and hate and know that all these feelings have a place in their world.
I am thus sympathetic to Walter Freeman's recent discussion of "A Neurobiological Role of Music in Social Bonding." Freeman is particularly interested in those rituals where a core group of celebrants move from one status in society to another, as from child to adult, or single to married. In these situations the bonds between individuals must be altered in deep and fundamental ways, ways requiring considerable rewiring in the core and autobiographical selves. Freeman suggests that such rituals involve a neuropeptide called oxytocin--which, I mention for those of you who are psychoanalytically inclined, is also released during orgasm and during lactation. Freeman (2000, 418) asserts that oxytocin "appears to act by dissolving preexisting learning by loosening the synaptic connections in which prior knowledge is held. This opens an opportunity for learning new knowledge. The meltdown does not instill knowledge. It clears the path for the acquisition of new understanding through behavioral actions that are shared with others" (cf. Freeman 1995, 120ff.). As the oxytocin-saturated individuals are moving to the rhythms of well-established ritual, or, for that matter, are moved by actors playing Antony and Cleopatra on the state, their synaptic connections are restructured in patterns guided and influenced by the events in the ritual, or the words in the text.
To be sure, we do not all respond in the same way, our synapses are not reorganized into the same patterns. What is important is that we subject ourselves to the same ritual process, the same patterns of words in texts, paint on canvas, rhythms in music and dance. In the larger orbits of our social interactions, these common rituals keep us in within range of one another so that we can negotiate deep understandings when the need arises. These rituals provide a space in which we can mold our selves to one another as the infant molds her actions to those of her mother. On those occasions when the gods stick their tongues out at us we all blink our eyes and respond in kind.
10. Conclusion: Beyond the Ghost
What then, has become of the ghostly self in the neural machine? Deep within that machine, we have Damasio's proto-self and within that, the reticular formation. While the RF may seem like a master controller, it isn't, not quite. The RF exerts global control within the brain and thereby commits the organism to a particular behavioral mode. But it doesn't control what the organism does when it is in that mode. One may be in eating mode, but decisions about just what and how you will eat are not made in the RF, which simply does not have a sufficiently fine-grained apprehension of the world to make such decisions. Those decisions reside with other brain structures. And so it goes with the other modes, exploration, mating, sleep, etc. Fundamental as it is, the proto-self does not run the show. It is not the fabled ghost in the neural machine.
Nor are we going to find a convincing ghost in the ego/alter mechanism that pulls the strings on personal pronouns. That mechanism plays a critical role in managing conversation and in facilitating empathetic identification with others, both real and imaginary. But it does not control the mind; it is but a piece of the mind's machinery.
However, it is this bit of machinery that is responsible for the illusion of a self. By supporting self-reference it is central to the manipulation of one's self image. It is this bit of machinery that allows us to assert that body, brain, mind, are all mine, thereby creating the illusion of a singular self running through all. Beyond that, this machinery is responsible for one of the more vexing philosopher's illusions. Recall the infinite regress we encountered while considering the image on the box of the COSMIC RAY GUN. The recursive process that allowed us to project an endless series of boxes on boxes can do the same thing when applied to "the self." You begin, in the manner of the eternal sophomore, by positing an Observed Self and an Observing Self. The same mechanism that allowed us to imagine just one more Raygun boy on just one more COSMIC RAY GUN box, will allow us to treat the Observing Self as an object of observation by another Observing Self. We then repeat the maneuver until we become bored with it. This infinite regress is silly, and the succession of selves it engenders is without significant content. They are illusory. Now that we have a plausible account of how neural machinery can construct that ghost, we need not take the ghost at face value.
The fact that we have exorcised the ghostly self does not, of course, mean that no self remains. The neural systems Damasio has examined under the rubric of the neural self certainly are real. But they do not constitute a single module in the brain nor do they exert executive control. They are widely distributed and have extensive connections with one another and with other brain regions. The personal pronoun system is but one circuit in the neural self, though its effects are far ranging and, in the limit, often the source of paradox. The recursive games one can play using such mechanisms are as abstract as any the human mind has conceived. By contrast, the neurochemical regulators of the proto-self are deep in the muck and mud of our phylogenetic history. It is in the interaction between these systems that we find the ongoing and ever changing Self.
This paper is the fruit of my first substantial intellectual work since the death of my beloved friend, colleague, and teacher, David G. Hays. As such, I would like to dedicate it to his memory. It is only fitting that this paper revisits the intellectual territory--cognitive network theory--we first explored together when I became his student in the middle 1970s. Richard Fritzson, another student of Hays, made very perceptive comments on an early draft, leading me to a substantially deeper technical exploration. I also wish to thank members of the International Paleopsychology Project, founded by Howard Bloom, for their comments on pieces of text that I posted online. Finally, I am indebted to PSYART's reviewers, whose trenchant yet encouraging comments forced me to think through a number of issues I'd allowed myself to side-step.
Notes
1On the notion of a neural state space, see Hobson (1999a, 1999b). Hobson's discussion of psychopathology in chapter eight of 1999a is particularly relevant, though it doesn't discuss dissociative identity disorder. One might also consider the notions of it mode-locking and intermittency in Kelso 1989, 114ff., 124ff.
As Hobson's state space is specifically about states of consciousness, this might be an appropriate place to make a remark or two about this model and consciousness, not by way of argument, but only for the purposes of stating my sense of things. I favor a distinction between basic consciousness and self-consciousness (cf. Edelman 1992). Though I have not given the matter a great deal of thought, I see no a priori reason not to grant some capacity for consciousness to all mammals, and perhaps to other vertebrates as well, but self-consciousness seems confined to humans and perhaps some apes as well--some apes are able to recognize themselves in a mirror (for a review, see Tomasselo and Call 1997, 331-337). Beyond this, I like the various accounts of consciousness advanced by Powers (1973), Tononi and Edelman (1998), and Freeman (1999). What these otherwise rather different models have in common is the notion that consciousness is not something that happens at a particular place in the brain; a conscious process can be organized at various places in the brain as the need arises. These models differ in how they characterize what it is that makes a process conscious. Those differences, however, are well beyond this discussion, which is about some few mechanisms and requirements of self consciousness. Return to main text
2 The term "neurofunctional area" is not a standard term in the neurosciences. I coined it to have one term that covers both functional regions of the cortex, which are generally called cortical regions or areas, and functional regions in subcortical structures, which are most frequently called nuclei. Return to main text
3 The model of the linguistic sign in Figure 2 is not the only one I have explored. Consider the figure below. We have schemas at both the sensorimotor (degree 2) and the systemic degree (degree 3), where systemic is understood as being the first purely cognitive degree in the mental system (Benzon and Hays 1988). The left side concerns meaning and indicates that the concept of dog has schemas in several NFAs. The visual NFA contains sensorimotor schemas representing the visual appearance of things while the haptic NFA contains sensorimotor schemas representing how things feel to the touch. There are, of course, other NFAs as well, olfactory, gustatory, kinesthetic, and so forth. There are intermodal systemic schemas which tie the mode-specific sensorimotor schemas together.
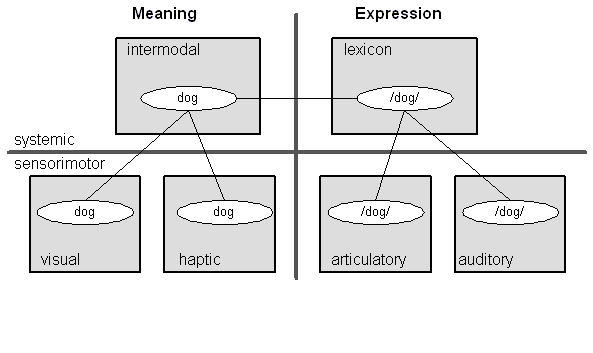
Language, the intermodal view
The right side of the diagram concerns linguistic expression. The auditory and articulatory NFAs are specifically linguistic, with one containing sensorimotor schemas representing the sounds of words and the other containing schemas for direct the utterance of words. (Obviously we have a general auditory NFA for non-linguistic sounds and other NFAs for non-linguistic motor control over the facial, neck, and trunk muscles which are also used in speaking.) The lexicon NFA contains systemic schemas which link the articulatory and auditory schemas together. Links between lexical schemas and intermodal schemas relate meanings to the sounds which express those meanings in language.
The model I have adopted in the main text (Figure 9) is a bit simpler. The intermodal NFA has been eliminated and lexical schemas are now directly connected to general sensorimotor schemas (visual, haptic, etc.) in addition to being connected to specifically articulatory and auditory schemas. One might well justify this on grounds of economy alone, for the inter-modal schemas would not, on the face of it, seem to carry any information about the represented object; they just serve as a gathering point for the mode-specific schemas. That function can easily be performed by the lexical node, thus eliminating the need for an inter-modal NFA.
However compelling that argument may be--and I'm quite sympathetic to it--conceptual economy comes into play only after empirical evidence runs out. In this case, there are empirical considerations. Older conceptions of cortical operation postulated the existence of large areas of intermodal tissue, with the amount increasing during the course of phylogeny. That conception was based more on conjecture than on actual anatomical evidence. As a consequence of more recent anatomical studies, however, Martin Sereno (1990) has questioned this notion, arguing that the amount of genuinely intermodal tissue is relatively slight even in advanced primates. Rather, we see an increase in the size and number of modality-specific areas. Sereno then goes on to argue against the notion that language is implemented in intermodal tissue and suggests that the lexicon is implemented in auditory tissue. While the argument I make in this essay does not depend on this point, I have nonetheless decided to follow Sereno in this conception. Return to main text
4 David Hays (1981, Benzon 1978) has proposed that the primary mechanism for the conditional control of behavior is something called an on-block: ON the detection of condition X, PERFORM action Y.
The brain has a variety of different NFAs involved in social interaction. It is thus a gross simplification to reduce that complexity to these two NFAs, one cortical (at the sensorimotor degree) and one subcortical. For more information on the neural substrates of social interaction see discussion in, e.g. Baron-Cohen 1995, MacLean 1990, Panksepp 1998. Return to main text
5 While this account may be a satisfactory way of approaching the cognitive grounding of first and second person pronouns, it still leaves us with third person pronouns. If ego and alter are derived from the infant's machinery for managing interaction with others, perhaps the third person pronouns have a similarly primitive derivation as well. It is not unreasonable to think that infants distinguish between persons in general and things. The third person animate pronouns (he, his, she, hers, etc.) would be derived from the primitive schema for person and apply to persons not currently engaging the infant in social interaction. The third person inanimate pronoun (it) would apply to any discernible object.
Note that I have an impression that David Hays and I once had a conversation in which he made remarks to this effect. I have no specific recollection of this conversation, and have no notes from it, but the possibility remains that this idea is one planted in my mind by David Hays. Return to main text
6 In fact all of the links in these diagrams should have such labels, for the links indicate the nature of the relationship between the schemas connected by them. I have not indicated the labels for the other links because a discussion of just what those labels mean would be distracting in the overall context of the argument. Note that Hays has suggested a scheme whereby link labels are not necessary (Hays 1981). Return to main text
7 Hays (1981) adopted Powers' model as his model of the sensorimotor degree in his cognitive model. This brings up the question of the nature of the relationship between Powers' stack of servomechanisms and these cognitive networks. Hays and I have given considerable attention to this problem, but I do not think we have gotten it quite right (Hays 1981, Benzon 1978). My current thinking is that the cognitive net stores schemas which can serve as reference levels for appropriate servos in the stacks--for there are multiple stacks operating in parallel. Return to main text
8 The general structure of the relationship between Piaget's stages and our cognitive system suggests that we need an episodic network to handle the older child's account of crawling (Benzon and Hays 1988). This is in addition to the sensorimotor and systemic networks we have been considering in this paper. Return to main text
9 Fauconnier's (1994) notion of mental spaces has some similarity to the concepts of episodic and gnomonic degrees (Bloom and Hays 1978; Hays 1981; Benzon 1978, Benzon and Hays 1988). Fauconnier's notion is explicitly that of a temporary construction which manages the meaning of linguistic strings and is distinct from ongoing and more or less permanent cognitive representations; such representations, may of course, enter into mental spaces. The discussions Hays and his students had about episodic and gnomonic degrees never really distinguished between their temporary "mental space" functions and their role as permanent cognitive structures. As a practical matter, most of our effort went into investigation the latter role, not the former.
In Hays' theory episodic structure does pretty much what Endel Tulving (1972) had in mind when he introduced the notion; it locates events in time and space: "On this time, in that place . . . " Episodic structure is thus close kin to Damasio's notion of the autobiographical self, though it is also a repository of general historical information unconnected with one's personal history. Gnomonic structure is a degree higher than episodic and Hays suggested that the purpose of gnomonic structure is to maintain the sense of reality. Gnomonic structure would thus mark an episodic space as being real or hypothetical in the sense that Lakoff (1996, 95ff.) discusses in his treatment of the self. For some explicit constructions using gnomonic structure in this way, see Benzon 1978, 187-196, 310ff., Benzon 1981, 258-262. Return to main text
10 The notion of recursion has played a critical role in modern linguistic theory. Recursive rules are central to the transformational component of Chomsky's early syntactic theory (cf. Chomsky 1957) and figure strongly in his argument against behaviorism (Chomsky 1959). However, a set of rules is just a collection of formal specifications. It cannot do anything by itself. If you want to do something, then you need some device to implement those rules. In Chomsky's methodology (1965, 3-15) the rules characterize linguistic competence, which is a different matter from the perceptual and motor procedures one uses in the actual performance of linguistic acts. In my view, the self structure is the mechanism that implements our recursive capacity.
I should also add that my views on language are quite different from Chomsky's in fundamental ways so I would not expect the self structure to be implementing Chomsky-style rules. However, Chomsky is the one who showed the relevance of recursion to the study of language and so I feel that I am explicating a notion of his, even if my explication is in quite a different style. Return to main text
11 William Powers (1973, 205-230) has developed models of memory and imagination that should be examined in this context. He presents no neural evidence in favor of his models, but the current neural observations follow directly from them. Return to main text
Amari, S, and Arbib, M. (1977). Competition and Cooperation in Neural Nets. In Systems Neuroscience. Jacqueline Metzler, ed. New York: Academic Press, 119-165.
Baron-Cohen, S. (1995). Mindblindness. Cambridge, MA: MIT Press.
Barr, Avrom, and Edward. A. Feigenbaum, eds. (1981). The Handbook of Artificial Intelligence, Vol. 1. Los Altos, CA: William Kaufmann.
Bartlett, Frederic C. (1932). Remembering. Cambridge: Cambridge University Press.
Benveniste, Emile (1958/1971). De la subjectivité dans le langage. Journal de Psychologie 55, 267ff. Repr. Subjectivity in Language. Problems in General Linguistics. Coral Gables, FL: University of Miami Press, 223-230.
Benzon, William L. (1976a). Cognitive Networks and Literary Semantics. MLN, 91: 952-982.
----- (1976b). Comment on Powers' Letter. MLN 91: 1619-1624.
----- (1978). Cognitive Science and Literary Theory. Diss., SUNY at Buffalo.
----- (1981). Lust in Action: An Abstraction. Language and Style 14: 251-270.
----- (1993a). The Evolution of Narrative and the Self. Journal of Social and Evolutionary Systems, 16.2: 129-155.
----- (1993b). Stages in the Evolution of Music. Journal of Social and Evolutionary Systems, 16.3: 283-296.
----- (1996). Culture as an Evolutionary Arena. Journal of Social and Evolutionary Systems, 19.4: 321-362.
----- and David G. Hays (1988). Principles and Development of Natural Intelligence. Journal of Social and Biological Structures 11: 293-322.
Bloom, D. and D. G. Hays (1978). Designation in English. In Anaphora in Discourse. John V. Hinds, ed. Champaign, Ill.: Linguistic Research, 1-68.
Bower, T. G. R. (1977). A Primer of Infant Development. San Francisco: W.H. Freeman.
Bownds, M. Deric (1999). The Biology of Mind. Bethesda, MD: Fitzgerald Science Press.
Bowsher, D. (1973). Brain, Behavior and Evolution. Brain, Behavior and Evolution 8: 386-396.
Bremner J. D., Krystal J. H., Charney D.S., and Southwick S. M. (1996). Neural Mechanisms in Dissociative Amnesia for Childhood Abuse: Relevance to the Current Controversy Surrounding the `false memory syndrome'. American Journal of Psychiatry 153.7 (Suppl): 71-82.
Cahill, L. and McGaugh, J. L. (1996). Modulation of memory storage. Current Opinion in Neurobiology 6: 237-242.
Chomsky, Noam (1957). Syntactic Structures. The Hague: Mouton.
----- (1959). A Review of B. F. Skinner's Verbal Behavior. Language 35: 26-58.
----- (1965). Aspects of the Theory of Syntax. Cambridge MA: MIT Press.
Church, Joseph (1966). Language and the Construction of Reality. New York: Vintage Books.
Damasio, Antonio (1994). Descartes' Error: Emotion, Reason, and the Human Brain. New York: Avon Books.
----- (1999a). Commentary by Damasio. Neuro-Psychoanalysis 1: 38-40.
----- (1999b). The Feeling of What Happens. New York: Harcourt, Brace.
Deren, Maya (1953). Divine Horsemen: The Living Gods of Haiti. London, New York: Thames and Hudson.
Edelman, Gerald M. (1992). Bright Air, Brilliant Fire. New York: Basic Books.
Epstein, Helen (1987). Music Talks: Conversations with Musicians. New York: McGraw-Hill.
Erikson, Erik (1963). Identity, Youth, and Crisis. New York: W. W. Norton.
Ewert, J-P. (1974). The Neural Basis of Visually Guided Behavior. Scientific American, 230.3, 35-49.
Fantz, Robert L. (1961). The Origin of Form Perception. Repr. Psychobiology. J. L. McGaugh, N. M. Weinberger, and R. E. Whalen, eds. San Francisco: W. H. Freeman.
Fauconnier, Gilles (1994). Mental Spaces. Cambridge, UK: Cambridge University Press.
Fink, G. R. et al. (1996). Cerebral Representation of One's Own Past: Neural Networks Involved in Autobiographical Memory. Journal of Neuroscience 16: 4275-4282.
Freeman, Walter J. (1995). Societies of Brains: A Study in the Neuroscience of Love and Hate. Hillsdale, NJ: Lawrence Erlbaum.
----- (1999). How Brains Make Up Their Minds. London: Weidenfeld and Nicholson.
----- (2000). Neurobiological Role of Music in Social Bonding. In The Origins of Music. Nils L. Wallin, Björn Merker, and Steven Brown, eds. Cambridge, MA: MIT Press, WHAT PAGES?.
Freud, Sigmund (1960) The Ego and the Id. Trans. Joan Riviere. New York: W. W. Norton.
Friedson, Steven M. (1996). Dancing Prophets: Musical Experience in Tumbuka Healing. Chicago: University of Chicago Press.
Gibson, J. J. (1979). The Ecological Approach to Visual Perception. Boston: Houghton Mifflin Company.
Green, André (1999). Consilience and Rigour. Neuro-Psychoanalysis 1: 40-44.
Hays, David G. (1981). Cognitive Structures. New Haven: HRAF Press.
----- (1976). On Alienation: An Essay in the Psycholinguistics of Science. In Theories of Alienation. Geyer, R. R., & Scheitzer, D. R., eds. Leiden: Martinus Nijhoff, 169-187.
----- (1992). The Evolution of Expressive Culture. Journal of Social and Biological Structures 15: 187-215.
Hinde, Robert A. (1974). The Biological Bases of Human Social Behavior. New York: McGraw-Hill.
Hobson, J. Allan (1999a). Consciousness. New York: W. H. Freeman.
----- (1999b). The New Neuropsychology of Sleep: Implications for Psychoanalysis. Neuro-Psychoanalysis 1: 157-183.
Humphrey, N. K. (1970). What the Frog's Eye Tells the Monkey's Brain. Brain, Behavior, and Evolution 3: 324-337.
Jakobson, Roman (1960). Linguistics and Poetics. In Style in Language. Thomas Sebeok, ed. Cambridge, MA: MIT Press, 350-77.
James, William (1890). The Principles of Psychology. New York: Henry Holt.
Jerison, Harry J. (1976). Paleoneurology and the Evolution of Mind. Scientific American 234.1: 90-101.
Johnson, Mark (1987). The Body in the Mind. Chicago: University of Chicago Press.
Johnson, Mark H. (1997). Developmental Cognitive Neuroscience. Oxford: Blackwell.
Joseph, R. (1999). The Neurology of Traumatic `dissociative amnesia': Commentary and Literature Review. Child Abuse and Neglect 23.8: 715-27.
Kelso, J. A. Scott (1995). Dynamic Patterns. Cambridge, MA: MIT Press.
Kilmer, W. L. McCulloch, W. S. and Blum, J. (1969). A Model of the Vertebrate Central Command System. International Journal of Man-Machine Studies 1: 279-309.
Kosslyn, Stephen M., and Koenig, Olivier (1995). Wet Mind: The New Cognitive Neuroscience. New York: Free Press.
Lakoff, George (1996). Sorry, I'm Not Myself Today: The Metaphor System for Conceptualizing the Self. In Spaces, Worlds, and Grammar. Gilles Fauconnier and Eve Sweetser, eds.Chicago: University of Chicago Press.
----- and Johnson, Mark (1999). Philosophy in the Flesh. New York: Basic Books.
Lamb, Sydney M. (1998). Pathways of the Brain. Amsterdam: John Benjamins.
LeDoux, Joseph (1996). The Emotional Brain. New York: Simon and Schuster.
Lenneberg, Eric (1967). The Biological Foundations of Language. New York: John Wiley & Sons. Inc.
Lettvin, Jerome Y., Maturana, Humberto R., McCulloch, Warren S. and Pitts, W. H. (1959). What the Frog's Eye Tells the Frog's Brain. Proceedings of the IRE 47: 1940-1959.
Luria, A. R. (1959). The Directive Function of Speech in Development and Dissolution. Word 15: 341-352, 453-464.
-----. (1973). The Working Brain. New York: Basic Books.
Lyons, John (1977). Semantics. Cambridge: Cambridge University Press.
MacLean, Paul D. (1990). The Triune Brain in Evolution. New York: Plenum Press.
McGaugh, James L. (1995). Emotional Activation, Neuromodulatory Systems, and Memory. In Memory Distortion. Daniel L. Schachter, ed. Cambridge, MA: Harvard University Press, 255-273.
Meltzoff, Andrew N., and M. Keith Moore (1995). Infants' Understanding of People and Things: From Body Imitation to Folk Psychology. In The Body and the Self. José Luis Bermúdez, Anthony Marcel, and Naomi Eilan, eds. Cambridge, MA: MIT Press.
Moruzzi, G. and Magoun, H. W. (1949). Brain stem reticular formation and activation of the EEG. Electroencephalography and Clinical Neurophysiology 1: 455-473.
Neisser, Ulric (1976). Cognition and Reality. San Francisco: W. H. Freeman.
Panksepp, Jaak (1998). Affective Neuroscience. New York: Oxford University Press.
----- (1999). Emotions as Viewed by Psychoanalysis and Neuroscience: An Exercise in Consilience. Neuro-Psychoanalysis 1: 15-38.
Piaget, Jean (1962). Play, Dreams and Imitation in Childhood. New York: W. W. Norton.
----- (1976). The Grasp of Consciousness. Cambridge, MA: Harvard University Press.
Posner, Michael I. and Marcus E. Raichle Images of Mind. New York: Scientific American Library, 1994.
Powers, William T. (1973). Behavior: The Control of Perception. Chicago: Aldine.
----- (1976). On Benzon and Models. MLN 91: 1612-1619.
Rappaport, David (1971). Emotions and Memory. New York: International Universities Press.
Regier, Terry (1996). The Human Semantic Potential. Cambridge, Mass: The MIT Press.
Rouget, Gilbert (1985). Music and Trance. Chicago: University of Chicago Press.
Sarnat, Harvey B., and Martin G. Netsky (1974). Evolution of the Nervous System. New York: Oxford University Press.
Schachter, Daniel L. (1996). Searching for Memory: The Brain, the Mind, and the Past. New York: Basic Books.
Schreiber, Flora Rheta (1973). Sybil. New York: Henry Regnery.
Sereno, M. (1990). Language and the Primate Brain. CRL Newsletter. Center for Research in Language. University of California at San Diego. Vol. 4.4. (http://crl.ucsd.edu/newsletter/4-4/Article1.html). Accessed May 28, 1996.
Spiegel, D. (1995). Hypnosis and Suggestion. In Memory Distortion. Daniel L. Schachter, ed. Cambridge, MA: Harvard University Press, 129-149.
Stanislavski, Konstantin. (1948). An Actor Prepares. New York: Theatre Arts Books.
Stoller, Robert J. (1973). Splitting. New York: Dell Publishing.
Thigpen, Corbett H. and Cleckley, Harvey M. (1957). The 3 Faces of Eve. New York: McGraw-Hill.
Tomasello, Michael, and Josep Call (1997). Primate Cognition. New York: Oxford University Press.
Tononi, G. and Edelman, Gerald M. (1998). Consciousness and Complexity. Science 282: 1846-1851.
Tulving, Endel (1972). Episodic and Semantic Memory. In Organization of Memory. Endel Tulving and Wayne Donaldson, eds. New York: Academic Press.
Vanderwolf, C. H. and Robinson, T. E. (1981). Reticulo-cortical Activity and Behavior: A Critique of the Arousal Theory and a New Synthesis. Behavioral and Brain Sciences, 4: 459-576.
Vygotsky, L. S. (1962). Thought and Language. Cambridge, MA: The MIT Press.
Received: June 19, 2000, Published: August 21, 2000. Copyright © 2000 William L. Benzon